
Gli amminoacidi sono mattoni fondamentali per la sintesi delle proteine in tutti gli organismi. Nelle piante, gli aminoacidi liberi "proteinogenici" svolgono ruoli aggiuntivi nell'assimilazione e nel trasporto dell'azoto, come composti di segnalazione, come osmoliti e come precursori per la produzione di vari ormoni, cofattori e altri composti importanti come la clorofilla. Le piante utilizzano gli aminoacidi anche per produrre collettivamente migliaia di composti specializzati che facilitano le interazioni ecologiche e forniscono risposte adattive agli stress ambientali.
I trasportatori localizzati sulla membrana plasmatica coinvolti nell'assorbimento degli aminoacidi da parte delle radici appartengono a tre famiglie all'interno della superfamiglia dei trasportatori di aminoacidi-poliamino-colina (APC): (i) le Amino Acid Permeases (AAPs), che funzionano come trasportatori di ampi substrati con affinità moderata; (ii) i Lysine/Histidine-like Transporters (LHTs), che importano aminoacidi neutri e acidi; e (iii) i Proline and Glycine Betaine Transporters (ProTs), che trasportano gli osmoliti prolina e glicina betaina (Tegeder e Masclaux-Daubresse, 2018).

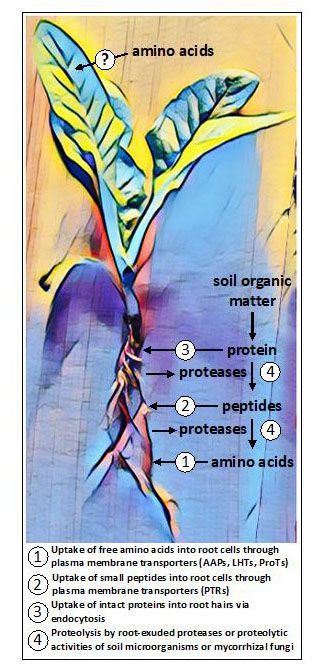
Figura 1. Potenziali vie di assorbimento degli aminoacidi riscontrate collettivamente nelle piante. Nelle radici, gli aminoacidi possono essere assunti in forma libera (monomerica) o in forma polimerica come piccoli peptidi o proteine. Alcune piante rilasciano proteasi per convertire le proteine e i peptidi presenti nel terreno in aminoacidi liberi, mentre altre piante si affidano alle attività dei funghi simbionti per la proteolisi e l'assorbimento. Le prove emergenti indicano che alcune piante possono essere in grado di acquisire aminoacidi applicati esogenamente ai tessuti aerei, anche se le vie di assorbimento presunte sono sconosciute.
Solo recentemente è stato scoperto che le radici di diverse piante non micorriziche trasudano enzimi proteolitici appartenenti alla famiglia delle cisteina-proteasi (Adamczyk et al., 2010) in grado di scindere le proteine sulla superficie della radice e nell'apoplasto della corteccia radicale (Paungfoo-Lonhienne et al., 2008). I piccoli peptidi e gli aminoacidi liberi rilasciati sono quindi disponibili per l'assorbimento da parte di trasportatori localizzati sulla membrana plasmatica. Le membrane delle cellule vegetali contengono anche chinasi recettoriali ricche di leucina (LRR-RK) che percepiscono ligandi peptidici e proteici (Hohmann et al., 2017). Questi recettori costituiscono il primo strato del sistema immunitario delle piante e i membri espressi nelle radici contribuiscono a controllare la formazione di barriere di diffusione nella fascia di Casparian (Okuda et al., 2020). Le proteine intatte sono anche in grado di entrare nei peli delle radici per endocitosi e vengono poi digerite all'interno delle cellule vegetali (Paungfoo-Lonhienne et al., 2008).
Nel suolo, le piante sono facilmente superate dai microrganismi per l'azoto organico. I microbi hanno tassi di ricambio più rapidi e rapporti superficie/volume più ampi, che consentono loro un maggiore accesso ai substrati nel suolo. Ciò fa sorgere dubbi sulla misura in cui aminoacidi, peptidi e proteine fungono da fonti significative di azoto per le piante (Näsholm et al., 2009). Poiché gli aminoacidi, i peptidi e le proteine applicati esogenamente hanno effetti benefici (Colla et al., 2017b) che vanno oltre il metabolismo dell'azoto, ciò potrebbe indicare altri ruoli per gli aminoacidi e i peptidi assunti dal suolo.
E altri tipi di piante? Studi di tracciatura con aminoacidi marcati con 15Nhanno dimostrato che le foglie di bentgrass strisciante (Stiegler et al., 2013) e di pesco (Furuya e Umemiya, 2002) sono in grado di assorbire aminoacidi attraverso il fogliame. Un recente studio di McCoy et al. (McCoy et al., 2020) ha inoltre dimostrato che il glutammato con doppia marcatura 15N,13C, applicato esogenamente al fogliame della gramigna, viene incorporato in prolina e acido γ-aminobutirrico (GABA). Questo lavoro è significativo perché suggerisce che la mineralizzazione sulla superficie fogliare è un destino minore per gli aminoacidi applicati e dimostra che gli aminoacidi forniti esogenamente possono essere incorporati nel metabolismo delle piante.



© BIOSTIMULANT.COM
TUTTI I DIRITTI RISERVATI