
Los aminoácidos son componentes vitales para la síntesis de proteínas en todos los organismos. En las plantas, los aminoácidos "proteinogénicos" libres desempeñan funciones adicionales en la asimilación y el transporte de nitrógeno, como compuestos de señalización, como osmolitos y como precursores para fabricar diversas hormonas, cofactores y otros compuestos importantes como la clorofila. Las plantas también utilizan aminoácidos para producir colectivamente miles de compuestos especializados que facilitan las interacciones ecológicas y proporcionan respuestas adaptativas al estrés ambiental.
Los transportadores localizados en la membrana plasmática que intervienen en la absorción de aminoácidos por las raíces pertenecen a tres familias de la superfamilia de transportadores de aminoácidos, poliamina y colina (APC): (i) las permeasas de aminoácidos (AAPs), que funcionan como transportadores de sustratos amplios con afinidades moderadas; (ii) los transportadores similares a la lisina/histidina (LHTs), que importan aminoácidos neutros y ácidos; y (iii) los transportadores de prolina y glicina betaína (ProTs), que transportan los osmolitos prolina y glicina betaína (Tegeder y Masclaux-Daubresse, 2018).

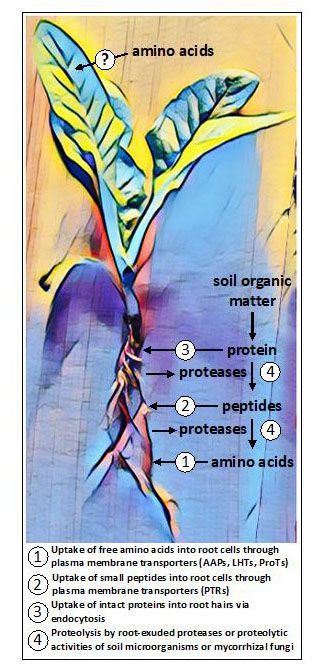
Figura 1. Vías potenciales de absorción de aminoácidos que se encuentran colectivamente en las plantas. En las raíces, los aminoácidos pueden ser absorbidos en sus formas libres (monoméricas) o en formas poliméricas como pequeños péptidos o proteínas. Algunas plantas liberan proteasas para convertir las proteínas y los péptidos del suelo en aminoácidos libres, mientras que otras dependen de las actividades de hongos simbiontes para la proteólisis y la absorción. Cada vez hay más pruebas que indican que algunas plantas pueden adquirir aminoácidos aplicados de forma exógena a los tejidos aéreos, aunque se desconocen las posibles vías de absorción.
Recientemente se ha descubierto que las raíces de varias plantas no micorrícicas exudan enzimas proteolíticas pertenecientes a la familia de las cisteína proteasas (Adamczyk et al., 2010) que pueden descomponer proteínas en la superficie de la raíz y en el apoplasto de la corteza radicular (Paungfoo-Lonhienne et al., 2008). Los pequeños péptidos liberados y los aminoácidos libres están entonces disponibles para ser absorbidos por transportadores localizados en la membrana plasmática. Las membranas de las células vegetales también contienen receptores quinasa ricos en leucina (LRR-RK) que detectan ligandos peptídicos y proteicos (Hohmann et al., 2017). Estos receptores forman la primera capa del sistema inmunitario de las plantas y los miembros expresados en las raíces ayudan a controlar la formación de barreras de difusión en la franja de Casparian (Okuda et al., 2020). Al parecer, las proteínas intactas también pueden entrar en los pelos radiculares por endocitosis y luego son digeridas dentro de las células vegetales (Paungfoo-Lonhienne et al., 2008).
En el suelo, los microorganismos superan fácilmente a las plantas en la competencia por el nitrógeno orgánico. Los microbios tienen tasas de renovación más rápidas y una mayor relación superficie/volumen, lo que les da un mayor acceso a los sustratos del suelo. Esto plantea la cuestión de hasta qué punto los aminoácidos, los péptidos y las proteínas son fuentes importantes de nitrógeno para las plantas (Näsholm et al., 2009). Dado que los aminoácidos, péptidos y proteínas aplicados exógenamente tienen efectos beneficiosos (Colla et al., 2017b) más allá del metabolismo del nitrógeno, esto puede apuntar a otras funciones de los aminoácidos y péptidos tomados del suelo.
¿Qué ocurre con otros tipos de plantas? Los estudios de rastreo con aminoácidos marcados con 15N demostraron que las hojas de creeping bentgrass (Stiegler et al., 2013) y melocotonero (Furuya y Umemiya, 2002) son capaces de absorber aminoácidos a través de su follaje. Un estudio reciente de McCoy et al. (McCoy et al., 2020) demostró además que el glutamato doblemente etiquetado con 15N,13Caplicado exógenamente al follaje del creeping bentgrass se incorpora a la prolina y al ácido γ-aminobutírico (GABA). Este trabajo es significativo porque sugiere que la mineralización en la superficie de la hoja es un destino menor para los aminoácidos aplicados y demuestra que los aminoácidos suministrados exógenamente pueden incorporarse al metabolismo de la planta.



© BIOSTIMULANT.COM
TODOS LOS DERECHOS RESERVADOS