
Аминокислоты являются жизненно важными строительными блоками для синтеза белков во всех организмах. В растениях свободные "протеиногенные" аминокислоты играют дополнительную роль в усвоении и переносе азота, в качестве сигнальных соединений, осмолитов и предшественников для производства различных гормонов, кофакторов и других основных соединений, таких как хлорофилл. Растения также используют аминокислоты для коллективного производства тысяч специализированных соединений, которые облегчают экологические взаимодействия и обеспечивают адаптивную реакцию на стрессы окружающей среды.
Локализованные на плазматической мембране транспортеры, участвующие в поглощении аминокислот корнями, принадлежат к трем семействам в рамках суперсемейства транспортеров аминокислот-полиамин-холина (APC): (i) аминокислотные пермеазы (AAPs), которые функционируют как транспортеры широких субстратов с умеренным сродством; (ii) лизин/гистидиноподобные транспортеры (LHTs), которые импортируют нейтральные и кислые аминокислоты; и (iii) транспортеры пролина и глицин-бетаина (ProTs), которые переносят осмолиты пролин и глицин-бетаин (Tegeder and Masclaux-Daubresse, 2018).

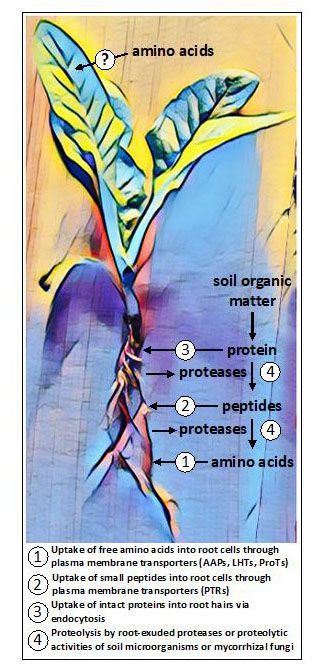
Рисунок 1. Потенциальные пути поглощения аминокислот, коллективно встречающиеся в растениях. В корнях аминокислоты могут поглощаться в свободной (мономерной) форме или в полимерной форме в виде небольших пептидов или белков. Некоторые растения выделяют протеазы для преобразования белков и пептидов в почве в свободные аминокислоты, в то время как другие растения полагаются на деятельность грибковых симбионтов для протеолиза и поглощения. Появляющиеся данные указывают на то, что некоторые растения могут приобретать аминокислоты, экзогенно вносимые в воздушные ткани, хотя предположительные пути поглощения неизвестны.
Совсем недавно было обнаружено, что корни нескольких различных немикоризных растений выделяют протеолитические ферменты, принадлежащие к семейству цистеиновых протеаз (Adamczyk et al., 2010), которые могут расщеплять белки на поверхности корня и в апопласте корневой коры (Paungfoo-Lonhienne et al., 2008). Высвобожденные малые пептиды и свободные аминокислоты затем доступны для поглощения транспортерами, локализованными в плазматической мембране. Мембраны растительных клеток также содержат рецепторные киназы с лейцин-богатым повтором (LRR-RK), которые воспринимают пептидные и белковые лиганды (Hohmann et al., 2017). Эти рецепторы образуют первый слой иммунной системы растений, а члены, экспрессированные в корнях, помогают контролировать формирование диффузионных барьеров в каспаровской полосе (Okuda et al., 2020). По имеющимся данным, неповрежденные белки также способны проникать в корневые волоски путем эндоцитоза и затем перевариваться внутри растительных клеток (Paungfoo-Lonhienne et al., 2008).
В почве растения легко конкурируют с микроорганизмами за органический азот. Микробы обладают более высокой скоростью оборота и большим отношением площади поверхности к объему, что дает им более широкий доступ к субстратам в почве. В связи с этим возникает вопрос о том, в какой степени аминокислоты, пептиды и белки служат значимыми источниками азота для растений (Näsholm et al., 2009). Поскольку экзогенно вносимые аминокислоты, пептиды и белки оказывают благотворное влияние (Colla et al., 2017b) не только на метаболизм азота, это может указывать на другие роли аминокислот и пептидов, взятых из почвы.
А как насчет других видов растений? Исследования с использованием 15N-меченыхаминокислот показали, что листья полевицы ползучей (Stiegler et al., 2013) и персикового дерева (Furuya and Umemiya, 2002) способны поглощать аминокислоты через листву. Недавнее исследование Маккоя и др. (McCoy et al., 2020) показало, что 15N,13Cглутамат с двойной меткой, экзогенно внесенный в листья полевицы, включается в пролин и γ-аминомасляную кислоту (ГАМК). Эта работа имеет большое значение, поскольку предполагает, что минерализация на поверхности листьев является незначительной судьбой для внесенных аминокислот, и демонстрирует, что экзогенно внесенные аминокислоты могут быть включены в метаболизм растений.



©2020 BIOSTIMULANT.COM
ВСЕ ПРАВА ЗАЩИЩЕНЫ