
氨基酸是所有生物体内合成蛋白质的重要组成部分。在植物中,游离的 "蛋白源 "氨基酸在氮的同化和运输中发挥着额外的作用,作为信号化合物,作为渗透压,以及作为制造各种激素、辅助因子和其他主要化合物如叶绿素的前体。植物还利用氨基酸集体生产成千上万的特殊化合物,以促进生态相互作用,并提供对环境压力的适应性反应。
参与根部氨基酸摄取的质膜定位转运体属于氨基酸-多胺-胆碱(APC)转运体超家族中的三个家族。(i) 氨基酸渗透酶(AAPs),作为具有中等亲和力的广泛底物转运体;(ii) 赖氨酸/组氨酸样转运体(LHTs),进口中性和酸性氨基酸;(iii) 脯氨酸和甘氨酸甜菜碱转运体(ProTs),运输渗透性物质脯氨酸和甘氨酸甜菜碱(Tegeder和Masclaux-Daubresse, 2018)。

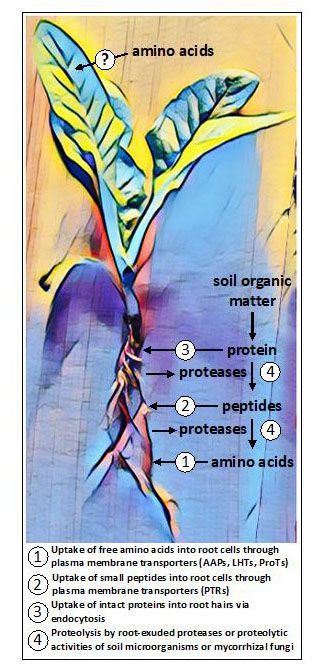
图1.在植物中集体发现的潜在的氨基酸摄取途径。在根部,氨基酸可以以其游离(单体)形式或以小肽或蛋白质的聚合形式被吸收。一些植物释放蛋白酶将土壤中的蛋白质和肽转化为游离氨基酸,而其他植物则依靠真菌共生体的活动进行蛋白质分解和摄取。新的证据表明,某些植物可能能够获得施加在气生组织上的外源性氨基酸,尽管推定的吸收途径尚不清楚。
最近才发现,几种不同的非根茎类植物的根部渗出属于半胱氨酸蛋白酶家族的蛋白质分解酶(Adamczyk等人,2010),可以分解根部表面和根部皮层的腹膜中的蛋白质(Paungfoo-Lonhienne等人,2008)。然后,释放的小肽和自由氨基酸可被质膜定位的运输工具吸收。植物细胞膜还含有富含亮氨酸的重复受体激酶(LRR-RKs),感知肽和蛋白质配体(Hohmann等人,2017)。这些受体形成植物免疫系统的第一层,在根部表达的成员有助于控制卡斯帕里安条带的扩散屏障的形成(Okuda等人,2020)。据报道,完整的蛋白质也能通过内吞作用进入根毛,然后在植物细胞内被消化(Paungfoo-Lonhienne等人,2008)。
在土壤中,植物很容易被微生物争夺有机氮。微生物有更快的周转率和更大的表面积与体积比,使它们更容易获得土壤中的底物。这就提出了氨基酸、肽和蛋白质作为植物氮的重要来源的程度问题(Näsholm等人,2009)。由于外源性的氨基酸、肽和蛋白质具有超出氮代谢的有益作用(Colla等人,2017b),这可能表明从土壤中吸收的氨基酸和肽的其他作用。
其他类型的植物呢?用15N标记的氨基酸进行的示踪研究表明,匍匐翦股颖(Stiegler等人,2013)和桃树(Furuya和Umemiya,2002)的叶片能够通过其叶片吸收氨基酸。麦考伊等人(McCoy et al., 2020)最近的一项研究进一步证明,15N,13C双标记的谷氨酸外施于匍匐茎叶,被纳入脯氨酸和γ-氨基丁酸(GABA)。这项工作很重要,因为它表明叶子表面的矿化作用是应用氨基酸的次要结果,并表明外源性氨基酸可以被纳入植物的新陈代谢中。


