
Amino acids are vital building blocks for synthesizing proteins in all organisms. In plants, free “proteinogenic” amino acids serve additional roles in the assimilation and transport of nitrogen, as signaling compounds, as osmolytes, and as precursors for making various hormones, cofactors, and other major compounds like chlorophyll. Plants also use amino acids to collectively produce thousands of specialized compounds that facilitate ecological interactions and provide adaptive responses to environmental stresses.
Plasma membrane-localized transporters involved in root amino acid uptake belong to three families within the Amino acid-Polyamine-Choline (APC) transporter superfamily: (i) the Amino Acid Permeases (AAPs), which function as broad substrate transporters with moderate affinities; (ii) the Lysine/Histidine-like Transporters (LHTs), which import neutral and acidic amino acids; and (iii) the Proline and Glycine Betaine Transporters (ProTs), which transport the osmolytes proline and glycine betaine (Tegeder and Masclaux-Daubresse, 2018).

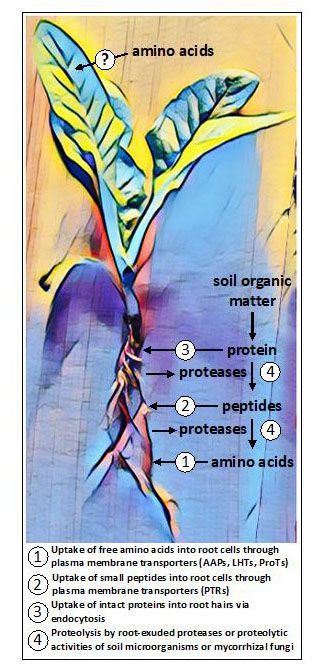
Figure 1. Potential routes of amino acid uptake collectively found in plants. In roots, amino acids can be taken up in their free (monomeric) forms or in polymeric forms as small peptides or proteins. Some plants release proteases to convert proteins and peptides in the soil into free amino acids, while other plants rely on the activities of fungal symbionts for proteolysis and uptake. Emerging evidence is indicating that certain plants may be able to acquire amino acids exogenously applied to aerial tissues, although the presumptive routes of uptake are unknown.
It was only more recently discovered that roots of several distinct nonmycorrhizal plants exude proteolytic enzymes belonging to the cysteine protease family (Adamczyk et al., 2010) that can break down proteins at the root surface and in the apoplast of the root cortex (Paungfoo-Lonhienne et al., 2008). The released small peptides and free amino acids are then available for uptake by plasma membrane-localized transporters. Plant cell membranes also contain leucine-rich repeat receptor kinases (LRR-RKs) that sense peptide and protein ligands (Hohmann et al., 2017). These receptors form the first layer of the plant immune system and members expressed in roots help to control formation of diffusion barriers in the Casparian strip (Okuda et al., 2020). Intact proteins are also reportedly able to enter root hairs by endocytosis and are then digested inside plant cells (Paungfoo-Lonhienne et al., 2008).
In the soil, plants are easily outcompeted by microorganisms for organic nitrogen. Microbes have faster turnover rates and larger surface area to volume ratios, giving them increased access to substrates in the soil. This then raises questions about the degree to which amino acids, peptides, and proteins serve as significant sources of nitrogen for plants (Näsholm et al., 2009). As exogenously applied amino acids, peptides, and proteins have beneficial effects (Colla et al., 2017b) beyond nitrogen metabolism, this may point to other roles for amino acids and peptides taken up from soil.
What about other types of plants? Tracer studies with 15N-labeled amino acids showed that creeping bentgrass (Stiegler et al., 2013) and peach tree (Furuya and Umemiya, 2002) leaves are capable of absorbing amino acids through their foliage. A recent study by McCoy et al. (McCoy et al., 2020) further demonstrated that 15N,13C double-labeled glutamate exogenously applied to creeping bentgrass foliage is incorporated into proline and γ-aminobutyric acid (GABA). This work is significant because it suggests that mineralization on the leaf surface is a minor fate for applied amino acids and demonstrates that exogenously supplied amino acids can be incorporated into plant metabolism.



©2020 BIOSTIMULANT.COM
ALL RIGHTS RESERVED