
Les acides aminés sont des éléments essentiels à la synthèse des protéines dans tous les organismes. Chez les plantes, les acides aminés libres "protéinogènes" jouent des rôles supplémentaires dans l'assimilation et le transport de l'azote, en tant que composés de signalisation, en tant qu'osmolytes et en tant que précurseurs pour la fabrication de diverses hormones, de cofacteurs et d'autres composés majeurs comme la chlorophylle. Les plantes utilisent également les acides aminés pour produire collectivement des milliers de composés spécialisés qui facilitent les interactions écologiques et fournissent des réponses adaptatives aux stress environnementaux.
Les transporteurs localisés dans la membrane plasmique impliqués dans l'absorption des acides aminés par les racines appartiennent à trois familles de la superfamille des transporteurs d'acides aminés, de polyamine et de choline (APC) : (i) les Amino Acid Permeases (AAPs), qui fonctionnent comme de larges transporteurs de substrat avec des affinités modérées ; (ii) les Lysine/Histidine-like Transporters (LHTs), qui importent les acides aminés neutres et acides ; et (iii) les Proline and Glycine Betaine Transporters (ProTs), qui transportent les osmolytes proline et glycine bétaïne (Tegeder et Masclaux-Daubresse, 2018).

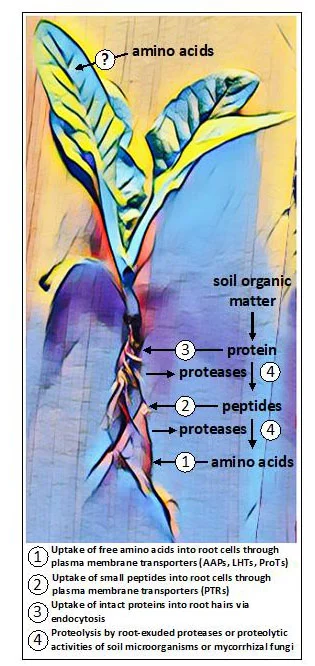
Figure 1. Voies potentielles d'absorption des acides aminés trouvées collectivement dans les plantes. Dans les racines, les acides aminés peuvent être absorbés sous leur forme libre (monomère) ou sous forme polymérique, comme de petits peptides ou protéines. Certaines plantes libèrent des protéases pour convertir les protéines et les peptides du sol en acides aminés libres, tandis que d'autres plantes dépendent des activités de symbiotes fongiques pour la protéolyse et l'absorption. De nouvelles preuves indiquent que certaines plantes peuvent être capables d'acquérir des acides aminés appliqués de manière exogène aux tissus aériens, bien que les voies présumées d'absorption soient inconnues.
Ce n'est que plus récemment que l'on a découvert que les racines de plusieurs plantes non mycorhiziennes distinctes exsudent des enzymes protéolytiques appartenant à la famille des cystéines protéases (Adamczyk et al., 2010) qui peuvent décomposer les protéines à la surface des racines et dans l'apoplaste du cortex racinaire (Paungfoo-Lonhienne et al., 2008). Les petits peptides et les acides aminés libres libérés sont alors disponibles pour être absorbés par des transporteurs localisés dans la membrane plasmique. Les membranes des cellules végétales contiennent également des récepteurs kinases à répétition riches en leucine (LRR-RK) qui détectent les ligands peptidiques et protéiques (Hohmann et al., 2017). Ces récepteurs forment la première couche du système immunitaire des plantes et les membres exprimés dans les racines aident à contrôler la formation de barrières de diffusion dans la bande caspienne (Okuda et al., 2020). Les protéines intactes seraient également capables de pénétrer dans les poils des racines par endocytose et sont ensuite digérées à l'intérieur des cellules végétales (Paungfoo-Lonhienne et al., 2008).
Dans le sol, les plantes sont facilement concurrencées par les micro-organismes pour l'azote organique. Les microbes ont des taux de renouvellement plus rapides et des rapports surface/volume plus importants, ce qui leur permet d'accéder plus facilement aux substrats dans le sol. On peut donc se demander dans quelle mesure les acides aminés, les peptides et les protéines constituent des sources importantes d'azote pour les plantes (Näsholm et al., 2009). Comme les acides aminés, les peptides et les protéines appliqués de manière exogène ont des effets bénéfiques (Colla et al., 2017b) au-delà du métabolisme de l'azote, cela peut indiquer d'autres rôles pour les acides aminés et les peptides prélevés dans le sol.
Qu'en est-il des autres types de plantes ? Des études de traçage avec des acides aminés marqués au 15Nont montré que les feuilles de l'agrostide rampante (Stiegler et al., 2013) et du pêcher (Furuya et Umemiya, 2002) sont capables d'absorber des acides aminés par leur feuillage. Une étude récente de McCoy et al. (McCoy et al., 2020) a en outre démontré que le glutamate doublement marqué 15N,13Cappliqué de manière exogène au feuillage de l'agrostide rampante est incorporé dans la proline et l'acide γ-aminobutyrique (GABA). Ce travail est important car il suggère que la minéralisation à la surface des feuilles est un destin mineur pour les acides aminés appliqués et démontre que les acides aminés fournis de manière exogène peuvent être incorporés dans le métabolisme des plantes.



©2020 BIOSTIMULANT.COM
TOUS DROITS RÉSERVÉS