
L'aumento della produttività delle colture, applicando al contempo pratiche sostenibili, è la sfida principale dell'agricoltura moderna per soddisfare le richieste di una popolazione mondiale in continua crescita che necessita di una maggiore qualità e sicurezza alimentare. A partire dalla Rivoluzione Verde, la qualità e la resa delle produzioni agricole sono state migliorate grazie all'applicazione di fertilizzanti chimici e pesticidi, con un impatto sulla sostenibilità economica e ambientale.
Tuttavia, negli ultimi decenni, la produttività agricola ha raggiunto una fase di stagnazione in cui le rese non sono aumentate proporzionalmente alla quantità di fertilizzanti applicati, evidenziando la necessità di migliorare l'efficienza di utilizzo delle risorse da parte delle colture (Pii et al., 2015a). Quando si considera la nutrizione minerale delle piante, la ricerca deve concentrarsi su un volume specifico del suolo, la rizosfera, caratterizzata da interazioni molto strette tra radici, microrganismi e il suolo stesso (Hinsinger et al., 2009). Infatti, le relazioni che si stabiliscono nella rizosfera giocano un ruolo centrale nel modulare la biodisponibilità dei nutrienti minerali. Le piante rilasciano essudati radicali, tra cui composti organici a basso e alto peso molecolare, che sono coinvolti, tra le altre funzioni, nella mobilizzazione dei nutrienti minerali dalle particelle del suolo (Vives-Peris et al., 2020). Tuttavia, i composti organici a basso peso molecolare possono anche funzionare come un forte chemioattrattore per i batteri del suolo che possono utilizzarli come fonte di carbonio facilmente accessibile (Hinsinger et al., 2009). La maggior parte dei batteri della rizosfera (rizobatteri) sono commensali che sfruttano gli essudati radicali come nutrimento.
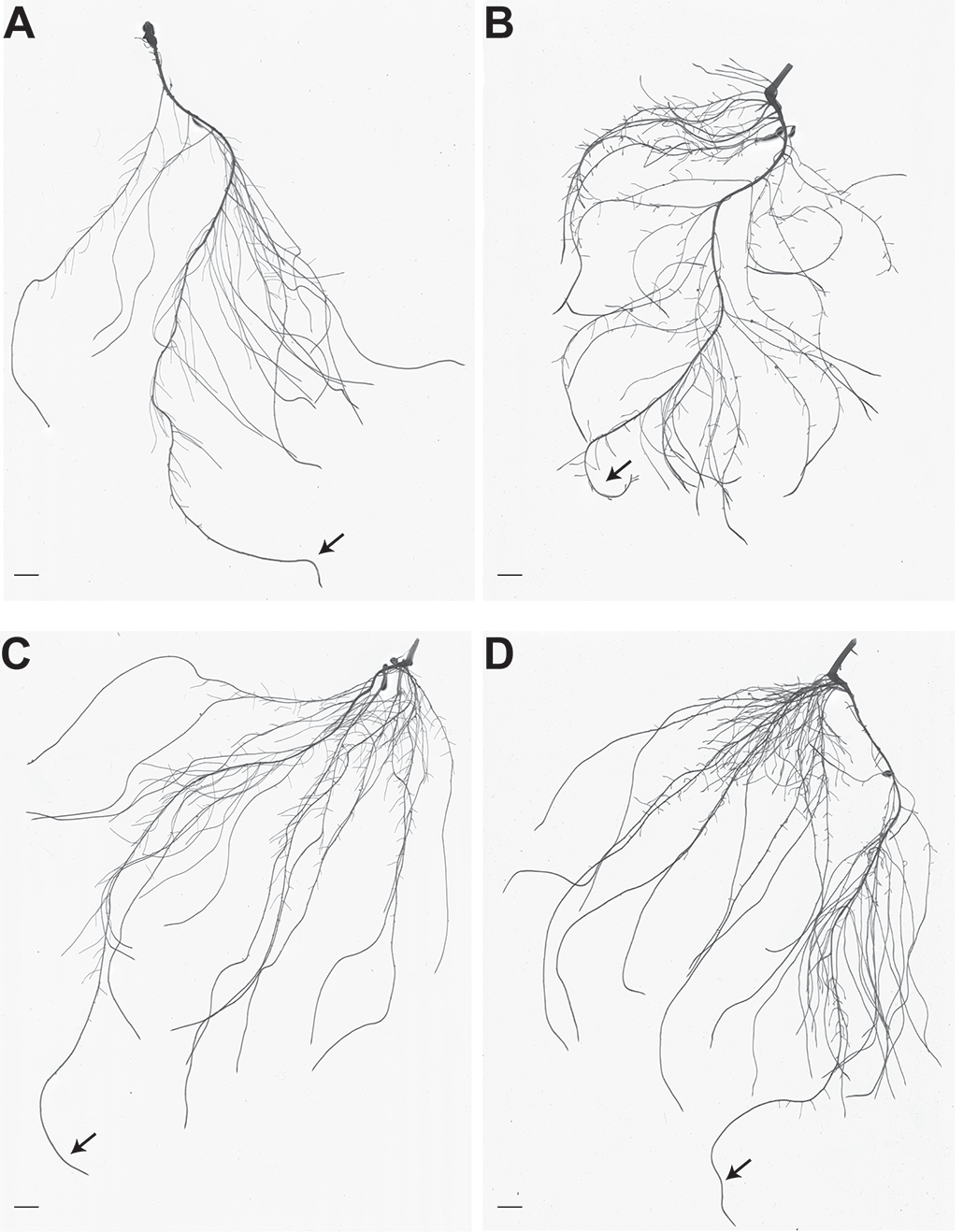
Figura 1. Architettura radicale di piante di cetriolo con e senza Fe, non inoculate o inoculate con A. brasilense. A. Immagine rappresentativa dell'apparato radicale di piante di cetriolo coltivate in una soluzione nutritiva con Fe sufficiente. B. Immagine rappresentativa dell'apparato radicale di piante di cetriolo coltivate in soluzione nutritiva povera di Fe. C. Immagine rappresentativa dell'apparato radicale di piante di cetriolo cresciute in soluzione nutritiva Fe-sufficiente e inoculate con A. brasilense. D. Immagine rappresentativa dell'apparato radicale di piante di cetriolo cresciute in soluzione nutritiva povera di Fe e inoculate con A. brasilense. Le scale delle barre rappresentano 1 cm. Le frecce indicano le radici primarie. (Figura modificata da Pii et al., 2016)
In conclusione, queste prove dimostrano che l'uso di PGPR in agricoltura potrebbe migliorare la capacità delle piante di sfruttare le risorse naturali già disponibili nel suolo, consentendo una riduzione dell'applicazione di fertilizzanti minerali e un aumento della sostenibilità dell'agricoltura. Inoltre, studi molto recenti hanno dimostrato che, oltre a migliorare la nutrizione minerale, l'uso di PGPR come biostimolanti vegetali può anche influenzare positivamente la qualità e i valori nutraceutici dei prodotti agricoli (Kolega et al., 2020; Pii et al., 2018).
Astolfi, S., Pii, Y., Mimmo, T., Lucini, L., Miras-moreno, M. B., Coppa, E., et al. (2020). La carenza singola e combinata di Fe e S modula in modo differenziato la composizione dell'essudato radicale nel pomodoro: Una doppia strategia per l'acquisizione del Fe? Int. J. Mol.Sci., 21(11), 4038. doi:10.3390/ijms21114038.
Brumbarova, T., Bauer, P. e Ivanov, R. (2015). Meccanismi molecolari che regolano l'assorbimento del ferro in Arabidopsis. Trends Plant Sci. 20, 124-133. doi:http://dx.doi.org/10.1016/j.tplants.2014.11.004.
Crecchio, C., Mimmo, T., Bulgarelli, D., Pertot, I., Pii, Y., Perazzolli, M., et al. (2018). "Microbioma del suolo benefico per la produzione agricola sostenibile", in, ed. E. Lichtfouse (Cham: Springer International Publishing), 443-481. doi:10.1007/978-3-319-94232-2_9.
de Santiago, A., García-López, A. M., Quintero, J. M., Avilés, M. e Delgado, A. (2013). Effetto del ceppo T34 di Trichoderma asperellum e dell'aggiunta di glucosio sulla nutrizione di ferro nel cetriolo coltivato su terreni calcarei. Soil Biol. Biochem. 57, 598-605. doi:10.1016/j.soilbio.2012.06.020.
Hinsinger, P., Bengough, A. G., Vetterlein, D. e Young, I. (2009). Rizosfera: biofisica, biogeochimica e rilevanza ecologica. Plant Soil 321, 117-152. doi:10.1007/s11104-008-9885-9.
Kobayashi, T. e Nishizawa, N. K. (2012). Assunzione, traslocazione e regolazione del ferro nelle piante superiori. Annu.Rev. Plant Biol. 63, 131-52. doi:10.1146/annurev-arplant-042811-105522.
Kolega, S., Moreno, B. M., Buffagni, V., Lucini, L., Valentinuzzi, F., Maver, M., et al. (2020). Profili nutraceutici di due cultivar di basilico dolce coltivate per via idroponica influenzati dalla composizione della soluzione nutritiva e dall'inoculazione con Azospirillum brasilense. Front. Plant Sci. 11, 1683. doi:10.3389/FPLS.2020.596000.
Marastoni, L., Pii, Y., Maver, M., Valentinuzzi, F., Cesco, S. e Mimmo, T. (2019). Ruolo di Azospirillum brasilense nell'innesco di diversi enzimi Fe chelato reduttasi in piante di cetriolo sottoposte a carenza di nutrienti e tossicità. Plant Physiol. Biochem. 136, 118-126. doi:10.1016/j.plaphy.2019.01.013.
Marschner, H. e Römheld, V. (1994). Strategie delle piante per l'acquisizione del ferro. Plant Soil 165, 261-274. doi:10.1007/BF00008069.
Pii, Y., Graf, H., Valentinuzzi, F., Cesco, S. e Mimmo, T. (2018). Gli effetti dei rizobatteri promotori della crescita delle piante (PGPR) sulla crescita e sulla qualità delle fragole. Acta Hortic., 231-238. doi:10.17660/actahortic.2018.1217.29.
Pii, Y., Marastoni, L., Springeth, C., Fontanella, M. C., Beone, G. M., Cesco, S., et al. (2016). Modulazione del processo di acquisizione del Fe da parte di Azospirillum brasilense in piante di cetriolo. Environ. Exp. Bot. 130, 216-225. doi:10.1016/j.envexpbot.2016.06.011.
Pii, Y., Mimmo, T., Tomasi, N., Terzano, R., Cesco, S. e Crecchio, C. (2015a). Interazioni microbiche nella rizosfera: influenze benefiche dei rizobatteri promotori della crescita delle piante sul processo di acquisizione dei nutrienti. Una rassegna. Biol. Fertil.Soils 51, 403-415. doi:10.1007/s00374-015-0996-1.
Pii, Y., Penn, A., Terzano, R., Crecchio, C., Mimmo, T. e Cesco, S. (2015b). Le interazioni pianta-microrganismo-suolo influenzano la disponibilità di Fe nella rizosfera di piante di cetriolo. Plant Physiol. Biochem. 87, 45-52. doi:10.1016/j.plaphy.2014.12.014.
Vives-Peris, V., de Ollas, C., Gómez-Cadenas, A. e Pérez-Clemente, R. M. (2020). Essudati radicali: dalla pianta alla rizosfera e oltre. Plant Cell Rep. 39, 3-17. doi:10.1007/s00299-019-02447-5.
Zhang, H., Sun, Y., Xie, X., Kim, M.-S., Dowd, S. E. e Paré, P. W. (2009). Un batterio del suolo regola l'acquisizione di ferro da parte delle piante attraverso meccanismi inducibili alla carenza. Plant J. 58, 568-577. doi:10.1111/j.1365-313X.2009.03803.x.
Zhao, L., Wang, F., Zhang, Y. e Zhang, J. (2014). Coinvolgimento del ceppo T6 di Trichoderma asperellum nella regolazione dell'acquisizione del ferro nelle piante. J. Basic Microbiol. 54, S115-S124. doi:10.1002/jobm.201400148.


© BIOSTIMULANT.COM
TUTTI I DIRITTI RISERVATI