
Повышение урожайности сельскохозяйственных культур при применении устойчивых методов является главной задачей современного сельского хозяйства для удовлетворения запросов постоянно растущего населения планеты, нуждающегося в более высоком качестве продуктов питания и продовольственной безопасности. Начиная с "зеленой революции", качество и урожайность сельскохозяйственной продукции были улучшены за счет более широкого применения химических удобрений и пестицидов, что повлияло на экономическую и экологическую устойчивость.
Однако в последние десятилетия производительность сельского хозяйства достигла фазы стагнации, когда урожайность не увеличивалась пропорционально количеству вносимых удобрений, что подчеркивает необходимость повышения эффективности использования ресурсов сельскохозяйственными культурами (Pii et al., 2015a). Когда рассматривается вопрос о минеральном питании растений, исследования должны быть сосредоточены на конкретном объеме почвы - ризосфере, характеризующейся очень тесным взаимодействием между корнями, микроорганизмами и самой почвой (Hinsinger et al., 2009). Фактически, отношения, установленные в ризосфере, играют центральную роль в модуляции биодоступности минеральных питательных веществ. Растения выделяют корневые экссудаты, включающие низко- и высокомолекулярные органические соединения, которые, помимо прочих функций, участвуют в мобилизации минеральных питательных веществ из почвенных частиц (Vives-Peris et al., 2020). Однако низкомолекулярные органические соединения могут также функционировать как сильный хемоаттрактант для почвенных бактерий, которые могут использовать их в качестве легкодоступного источника углерода (Hinsinger et al., 2009). Большинство ризосферных бактерий (ризобактерий) являются комменсалами, которые используют корневые экссудаты в качестве питания.
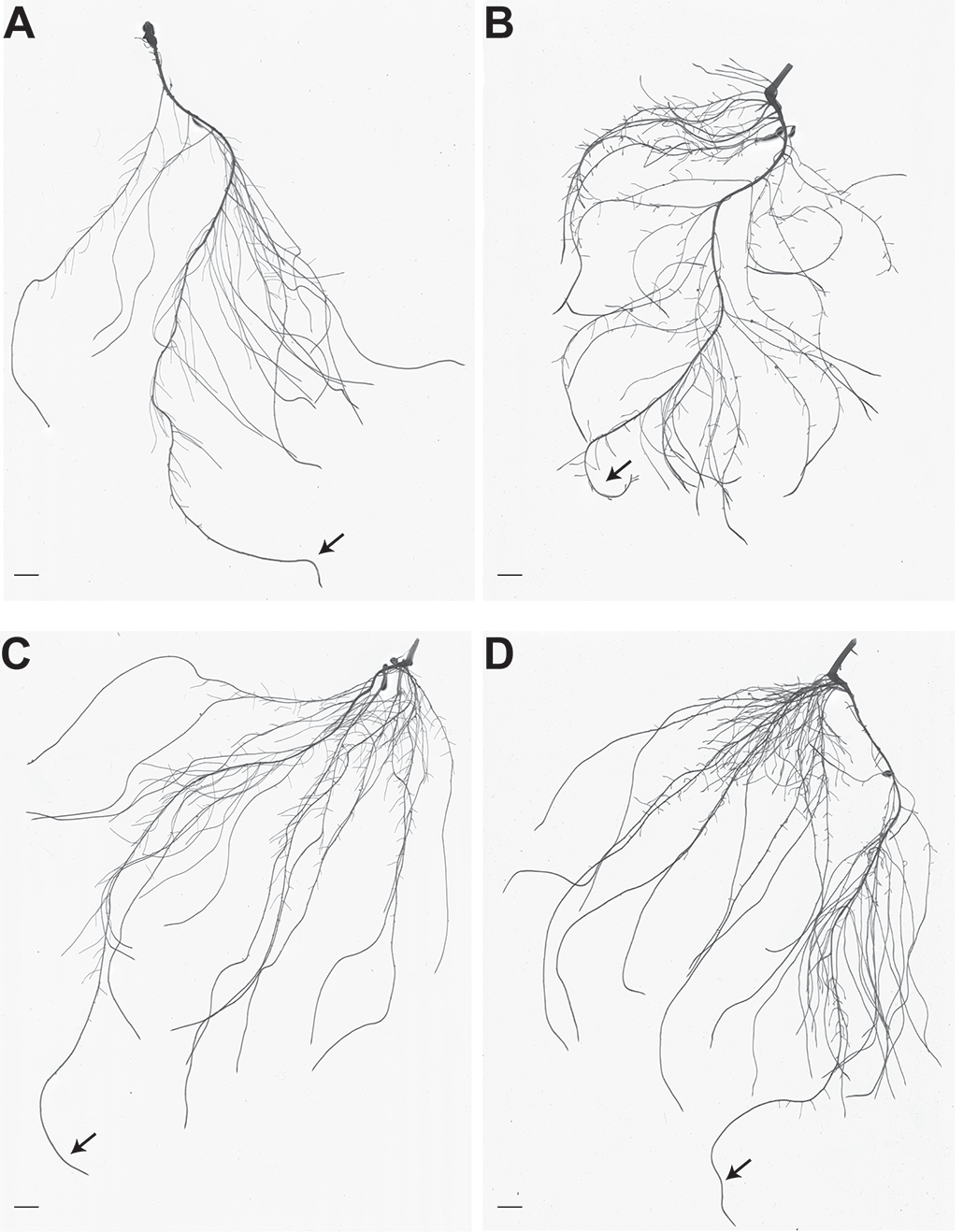
Рисунок 1. Архитектура корней растений огурца с достаточным содержанием Fe и дефицитом Fe, не инокулированных или инокулированных A. brasilense. A. Репрезентативная фотография корневой системы растений огурца, выращенных в питательном растворе с достаточным содержанием Fe. B. Репрезентативная фотография корневой системы огуречных растений, выращенных в питательном растворе с недостатком Fe. C. Репрезентативная фотография корневой системы огуречных растений, выращенных в питательном растворе с достаточным содержанием Fe и инокулированных A. brasilense. D. Репрезентативная фотография корневой системы растений огурца, выращенных в питательном растворе с недостатком Fe и инокулированных A. brasilense. Штриховые шкалы обозначают 1 см. Стрелки указывают на первичные корни. (Рисунок модифицирован из Pii et al., 2016)
В заключение, эти данные свидетельствуют о том, что использование PGPR в сельском хозяйстве может улучшить способность растений использовать природные ресурсы, уже имеющиеся в почве, что позволяет сократить применение минеральных удобрений и повысить устойчивость сельского хозяйства. Более того, совсем недавние исследования показали, что, помимо улучшения минерального питания, использование PGPR в качестве биостимуляторов растений может также положительно влиять на качество и нутрицевтическую ценность сельскохозяйственной продукции (Kolega et al., 2020; Pii et al., 2018).
Astolfi, S., Pii, Y., Mimmo, T., Lucini, L., Miras-moreno, M. B., Coppa, E., et al. (2020). Одиночный и комбинированный дефицит Fe и S дифференциально модулируют состав корневого экссудата у томата: Двойная стратегия для приобретения Fe? Int. J. Mol.Sci., 21(11), 4038. doi:10.3390/ijms21114038.
Брумбарова, Т., Бауэр, П., и Иванов, Р. (2015). Молекулярные механизмы, регулирующие поглощение железа арабидопсисом. Trends Plant Sci. 20, 124-133. doi:http://dx.doi.org/10.1016/j.tplants.2014.11.004.
Crecchio, C., Mimmo, T., Bulgarelli, D., Pertot, I., Pii, Y., Perazzolli, M., et al. (2018). "Полезный микробиом почвы для устойчивого сельскохозяйственного производства", под ред. E. Lichtfouse (Cham: Springer International Publishing), 443-481. doi:10.1007/978-3-319-94232-2_9.
де Сантьяго, А., Гарсия-Лопес, А. М., Кинтеро, Х. М., Авилес, М. и Дельгадо, А. (2013). Влияние штамма Trichoderma asperellum T34 и добавления глюкозы на питание железом огурцов, выращенных на известковых почвах. Почвенная биол. Biochem. 57, 598-605. doi:10.1016/j.soilbio.2012.06.020.
Hinsinger, P., Bengough, A. G., Vetterlein, D., and Young, I. (2009). Ризосфера: биофизика, биогеохимия и экологическая значимость. Plant Soil 321, 117-152. doi:10.1007/s11104-008-9885-9.
Кобаяси, Т., и Нишизава, Н. К. (2012). Поглощение, транслокация и регуляция железа у высших растений. Annu.Rev. Plant Biol. 63, 131-52. doi:10.1146/annurev-arplant-042811-105522.
Kolega, S., Moreno, B. M., Buffagni, V., Lucini, L., Valentinuzzi, F., Maver, M., et al. (2020). Нутрицевтические профили двух гидропонически выращенных сортов сладкого базилика в зависимости от состава питательного раствора и инокуляции Azospirillum brasilense. Front. Plant Sci. 11, 1683. doi:10.3389/FPLS.2020.596000.
Marastoni, L., Pii, Y., Maver, M., Valentinuzzi, F., Cesco, S., and Mimmo, T. (2019). Роль Azospirillum brasilense в запуске различных ферментов хелатной редуктазы Fe в растениях огурца, подверженных как дефициту питательных веществ, так и токсичности. Физиол. биохим. растений. 136, 118-126. doi:10.1016/j.plaphy.2019.01.013.
Маршнер, Х., и Рёмхельд, В. (1994). Стратегии растений для приобретения железа. Plant Soil 165, 261-274. doi:10.1007/BF00008069.
Pii, Y., Graf, H., Valentinuzzi, F., Cesco, S., and Mimmo, T. (2018). Влияние ризобактерий, стимулирующих рост растений (PGPR), на рост и качество клубники. Acta Hortic., 231-238. doi:10.17660/actahortic.2018.1217.29.
Pii, Y., Marastoni, L., Springeth, C., Fontanella, M. C., Beone, G. M., Cesco, S., et al. (2016). Модуляция процесса приобретения Fe с помощью Azospirillum brasilense в растениях огурца. Environ. Exp. Bot. 130, 216-225. doi:10.1016/j.envexpbot.2016.06.011.
Pii, Y., Mimmo, T., Tomasi, N., Terzano, R., Cesco, S., and Crecchio, C. (2015a). Микробные взаимодействия в ризосфере: благотворное влияние ризобактерий, стимулирующих рост растений, на процесс приобретения питательных веществ. A review. Biol. Fertil.Soils 51, 403-415. doi:10.1007/s00374-015-0996-1.
Pii, Y., Penn, A., Terzano, R., Crecchio, C., Mimmo, T., and Cesco, S. (2015b). Взаимодействие между растениями, микроорганизмами и почвой влияет на доступность Fe в ризосфере огуречных растений. Физиол. биохим. растений. 87, 45-52. doi:10.1016/j.plaphy.2014.12.014.
Vives-Peris, V., de Ollas, C., Gómez-Cadenas, A., and Pérez-Clemente, R. M. (2020). Корневые экссудаты: от растения к ризосфере и далее. Plant Cell Rep. 39, 3-17. doi:10.1007/s00299-019-02447-5.
Zhang, H., Sun, Y., Xie, X., Kim, M.-S., Dowd, S. E., and Paré, P. W. (2009). Почвенная бактерия регулирует приобретение железа растениями с помощью механизмов, вызывающих дефицит. Plant J. 58, 568-577. doi:10.1111/j.1365-313X.2009.03803.x.
Zhao, L., Wang, F., Zhang, Y., and Zhang, J. (2014). Вовлечение Trichoderma asperellum штамм T6 в регулирование приобретения железа в растениях. J. Basic Microbiol. 54, S115-S124. doi:10.1002/jobm.201400148.


© BIOSTIMULANT.COM
ВСЕ ПРАВА ЗАЩИЩЕНЫ