
Aumentar la productividad de los cultivos, aplicando al mismo tiempo prácticas sostenibles, es el principal reto de la agricultura moderna para satisfacer las demandas de una población mundial cada vez más numerosa y necesitada de alimentos de mayor calidad y seguridad alimentaria. A partir de la Revolución Verde, la calidad y el rendimiento de las producciones agrícolas se han mejorado mediante el aumento de la aplicación de fertilizantes químicos y plaguicidas, lo que ha repercutido en la sostenibilidad económica y medioambiental.
Sin embargo, a lo largo de las últimas décadas, la productividad agrícola ha alcanzado una fase de estancamiento en la que los rendimientos no han aumentado proporcionalmente a la cantidad de fertilizantes aplicados, lo que pone de manifiesto la necesidad de mejorar la eficiencia en el uso de los recursos por parte de los cultivos (Pii et al., 2015a). Cuando se considera la nutrición mineral de las plantas, la investigación debe centrarse en un volumen específico del suelo, la rizosfera, caracterizada por interacciones muy estrechas entre las raíces, los microorganismos y el propio suelo (Hinsinger et al., 2009). De hecho, las relaciones que se establecen en la rizosfera desempeñan un papel fundamental en la modulación de la biodisponibilidad de los nutrientes minerales. Las plantas liberan exudados radiculares, incluyendo compuestos orgánicos de bajo y alto peso molecular, que están implicados, entre otras funciones, en la movilización de nutrientes minerales desde las partículas del suelo (Vives-Peris et al., 2020). Sin embargo, los compuestos orgánicos de bajo peso molecular también pueden funcionar como un fuerte quimioatrayente para las bacterias del suelo que pueden utilizarlos como una fuente de carbono de fácil acceso (Hinsinger et al., 2009). La mayoría de las bacterias de la rizosfera (rizobacterias) son comensales que aprovechan los exudados de las raíces como alimento.
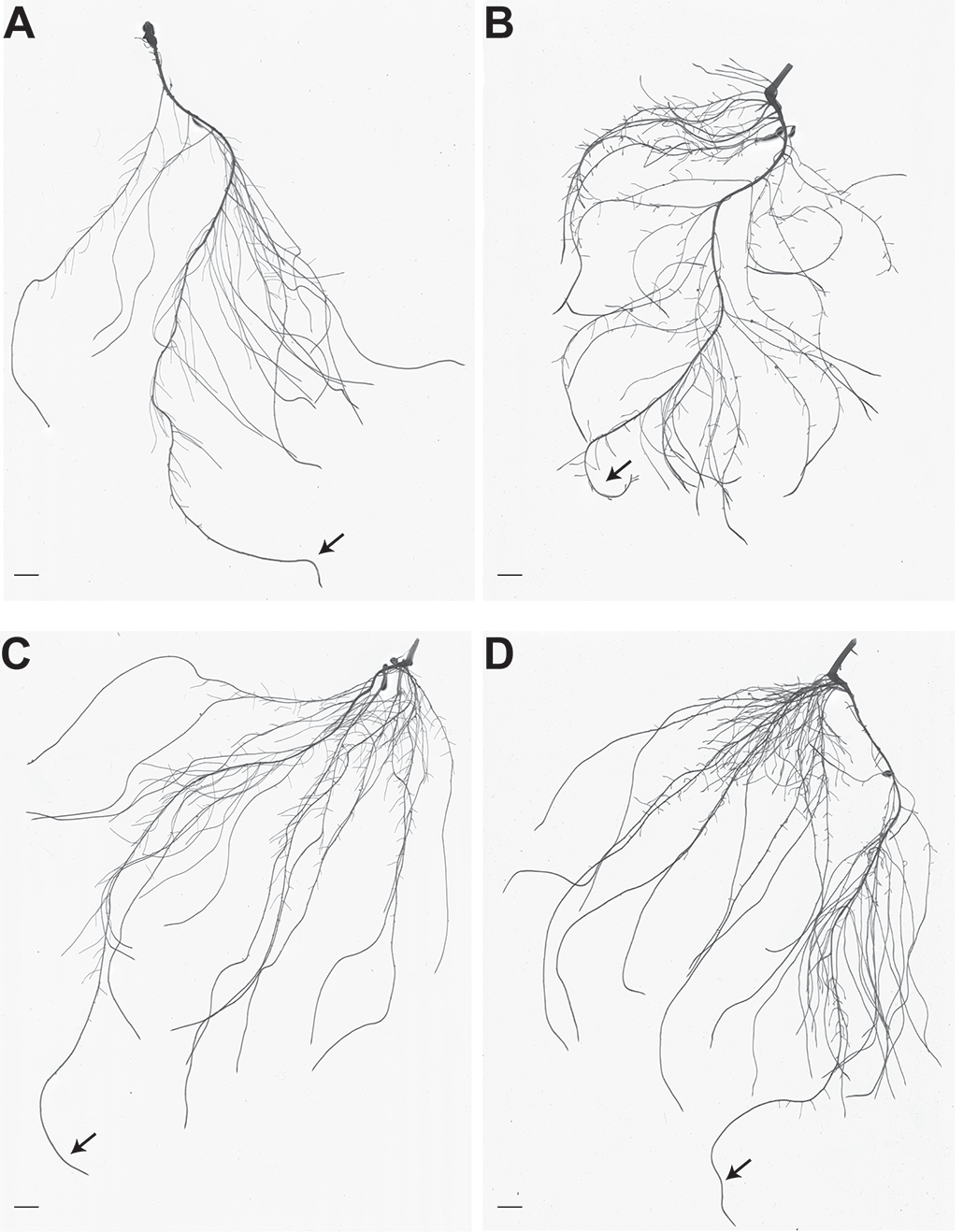
Figura 1. Arquitectura radicular de plantas de pepino con y sin Fe, inoculadas o no con A. brasilense. A . Imagen representativa del sistema radicular de plantas de pepino cultivadas en solución nutritiva con suficiente Fe. B . Imagen representativa del sistema radicular de plantas de pepino cultivadas en solución nutritiva deficiente en Fe. C . Imagen representativa del sistema radicular de plantas de pepino cultivadas en solución nutritiva con suficiente Fe e inoculadas con A. brasilense. D . Imagen representativa del sistema radicular de plantas de pepino cultivadas en solución nutritiva deficiente en Fe e inoculadas con A. brasilense. Las escalas de las barras representan 1 cm. Las flechas indican raíces primarias. (Figura modificada a partir de Pii et al., 2016)
En conclusión, estas evidencias demuestran que el uso de PGPR en agricultura podría mejorar la capacidad de las plantas para explotar los recursos naturales ya disponibles en el suelo, permitiendo una reducción en la aplicación de fertilizantes minerales y un aumento de la sostenibilidad de la agricultura. Además, estudios muy recientes han demostrado que, además de mejorar la nutrición mineral, el uso de PGPR como bioestimulantes vegetales también puede afectar positivamente a la calidad y a los valores nutracéuticos de los productos agrícolas (Kolega et al., 2020; Pii et al., 2018).
Astolfi, S., Pii, Y., Mimmo, T., Lucini, L., Miras-moreno, M. B., Coppa, E., et al. (2020). Single and Combined Fe and S Deficiency Differentially Modulate Root Exudate Composition in Tomato : A Double Strategy for Fe Acquisition ? Int. J. Mol.Sci., 21(11), 4038. doi:10.3390/ijms21114038.
Brumbarova, T., Bauer, P. e Ivanov, R. (2015). Mecanismos moleculares que rigen la absorción de hierro en Arabidopsis. Trends Plant Sci. 20, 124-133. doi:http://dx.doi.org/10.1016/j.tplants.2014.11.004.
Crecchio, C., Mimmo, T., Bulgarelli, D., Pertot, I., Pii, Y., Perazzolli, M., et al. (2018). "Microbioma beneficioso del suelo para la producción agrícola sostenible", en, ed. E. Lichtfouse (Cham: Springer International Publishing), 443-481. doi:10.1007/978-3-319-94232-2_9.
de Santiago, A., García-López, A. M., Quintero, J. M., Avilés, M., y Delgado, A. (2013). Efecto de la cepa T34 de Trichoderma asperellum y la adición de glucosa sobre la nutrición de hierro en pepino cultivado en suelos calcáreos. Biol. Biochem. 57, 598-605. doi:10.1016/j.soilbio.2012.06.020.
Hinsinger, P., Bengough, A. G., Vetterlein, D., y Young, I. (2009). Rizosfera: biofísica, biogeoquímica y relevancia ecológica. Plant Soil 321, 117-152. doi:10.1007/s11104-008-9885-9.
Kobayashi, T., y Nishizawa, N. K. (2012). Iron uptake, translocation, and regulation in higher plants. Annu.Rev. Plant Biol. 63, 131-52. doi:10.1146/annurev-arplant-042811-105522.
Kolega, S., Moreno, B. M., Buffagni, V., Lucini, L., Valentinuzzi, F., Maver, M., et al. (2020). Nutraceutical profiles of two hydroponically grown sweet basil cultivars as affected by the composition of the nutrient solution and the inoculation with Azospirillum brasilense. Front. Plant Sci. 11, 1683. doi:10.3389/FPLS.2020.596000.
Marastoni, L., Pii, Y., Maver, M., Valentinuzzi, F., Cesco, S., and Mimmo, T. (2019). Papel de Azospirillum brasilense en la activación de diferentes enzimas quelato reductasa de Fe en plantas de pepino sometidas tanto a deficiencia de nutrientes como a toxicidad. Plant Physiol Biochem. 136, 118-126. doi:10.1016/j.plaphy.2019.01.013.
Marschner, H., y Römheld, V. (1994). Estrategias de las plantas para la adquisición de hierro. Plant Soil 165, 261-274. doi:10.1007/BF00008069.
Pii, Y., Graf, H., Valentinuzzi, F., Cesco, S., y Mimmo, T. (2018). Los efectos de las rizobacterias promotoras del crecimiento vegetal (PGPR) en el crecimiento y la calidad de las fresas. Acta Hortic., 231-238. doi:10.17660/actahortic.2018.1217.29.
Pii, Y., Marastoni, L., Springeth, C., Fontanella, M. C., Beone, G. M., Cesco, S., et al. (2016). Modulación del proceso de adquisición de Fe por Azospirillum brasilense en plantas de pepino. Environ. Exp. Bot. 130, 216-225. doi:10.1016/j.envexpbot.2016.06.011.
Pii, Y., Mimmo, T., Tomasi, N., Terzano, R., Cesco, S. y Crecchio, C. (2015a). Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil.Soils 51, 403-415. doi:10.1007/s00374-015-0996-1.
Pii, Y., Penn, A., Terzano, R., Crecchio, C., Mimmo, T., y Cesco, S. (2015b). Plant-microorganism-soil interactions influence the Fe availability in the rhizosphere of cucumber plants. Plant Physiol. Biochem. 87, 45-52. doi:10.1016/j.plaphy.2014.12.014.
Vives-Peris, V., de Ollas, C., Gómez-Cadenas, A., y Pérez-Clemente, R. M. (2020). Exudados radiculares: de la planta a la rizosfera y más allá. Plant Cell Rep. 39, 3-17. doi:10.1007/s00299-019-02447-5.
Zhang, H., Sun, Y., Xie, X., Kim, M.-S., Dowd, S. E., y Paré, P. W. (2009). A soil bacterium regulates plant acquisition of iron via deficiency-inducible mechanisms. Plant J. 58, 568-577. doi:10.1111/j.1365-313X.2009.03803.x.
Zhao, L., Wang, F., Zhang, Y., y Zhang, J. (2014). Involvement of Trichoderma asperellum strain T6 in regulating iron acquisition in plants. J. Basic Microbiol. 54, S115-S124. doi:10.1002/jobm.201400148.


© BIOSTIMULANT.COM
TODOS LOS DERECHOS RESERVADOS