
An increase in crop productivity, while applying sustainable practices, is the main challenge for modern agriculture to satisfy the requests of the ever-growing world population in need of higher food quality and food security. Starting with the Green Revolution, the quality and the yields of agricultural productions have been improved by enhancing the application of chemical fertilizers and pesticides, impacting economic and environmental sustainability.
However, through the last decades, agricultural productivity has reached a stagnation phase in which yields have not proportionally increased with the amount of fertilizers applied, highlighting the need for improving resources use efficiency by crops (Pii et al., 2015a). When plants’ mineral nutrition is considered, research needs to focus on a specific soil volume, the rhizosphere – characterized by very close interactions between roots, microorganisms, and the soil itself (Hinsinger et al., 2009). In fact, the relationships established in the rhizosphere play a central role in modulating the bioavailability of mineral nutrients. Plants release root exudates, including low and high molecular weight organic compounds, that are involved, among other functions, in the mobilization of mineral nutrients from soil particles (Vives-Peris et al., 2020). However, low molecular weight organic compounds can also function as a strong chemoattractant for soil bacteria that can use them as an easily accessible carbon source (Hinsinger et al., 2009). The majority of rhizosphere bacteria (rhizobacteria) are commensals that take advantage of root exudates as nourishment.
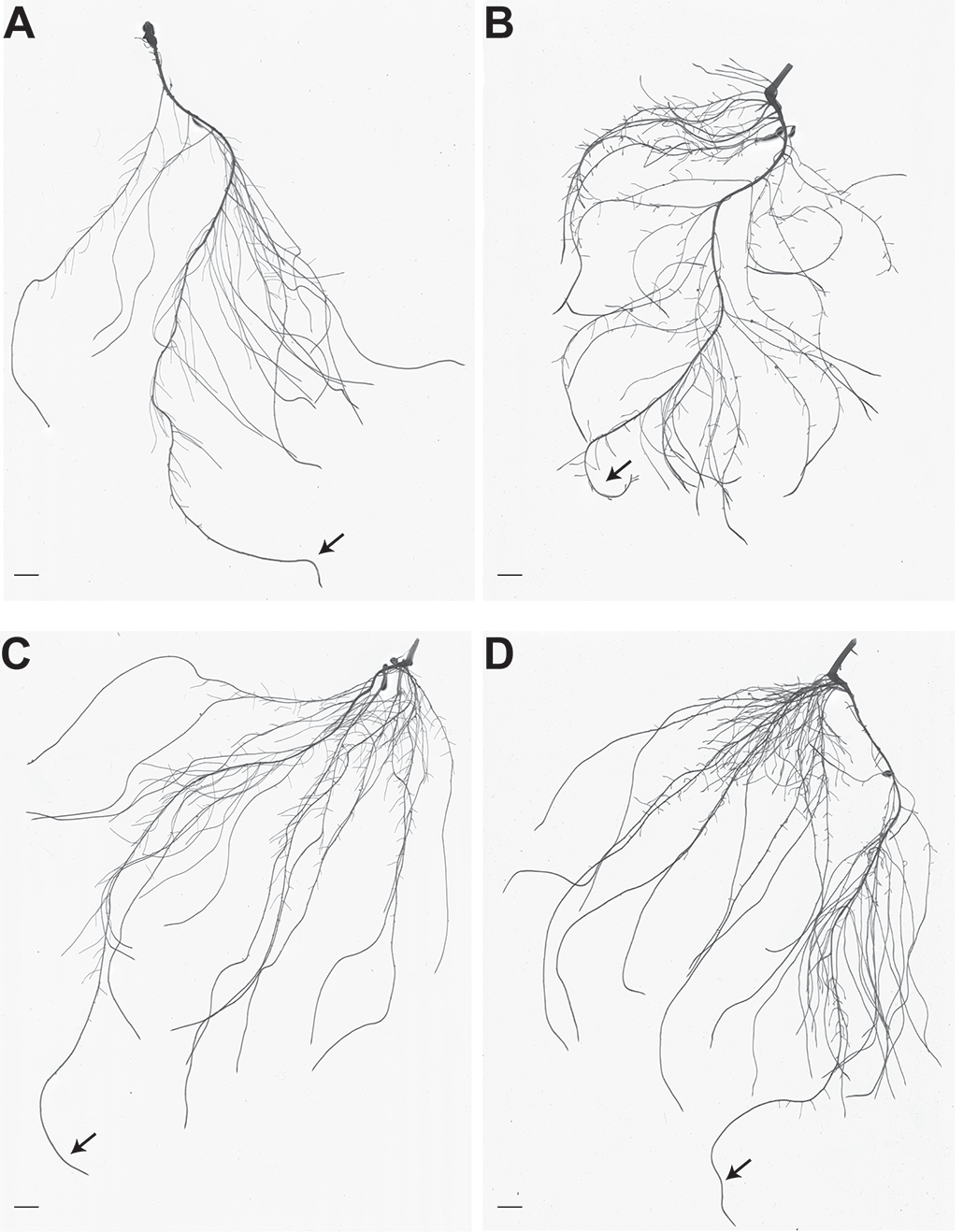
Figure 1. Root architecture of Fe sufficient and Fe deficient cucumber plants, either non-inoculated or inoculated with A. brasilense. A. Representative picture of the root system of cucumber plants grown in Fe-sufficient nutrient solution. B. Representative picture of the root system of cucumber plants grown in Fe-deficient nutrient solution. C. Representative picture of the root system of cucumber plants grown in Fe-sufficient nutrient solution and inoculated with A. brasilense. D. Representative picture of the root system of cucumber plants grown in Fe-deficient nutrient solution and inoculated with A. brasilense. Bar scales represent 1 cm. Arrows indicate primary roots. (Figure modified from Pii et al., 2016)
In conclusion, these pieces of evidence demonstrate that the use of PGPR in agriculture might improve the ability of plants to exploit the natural resources already available in the soil, allowing a reduction in the application of mineral fertilizers and an increase in agriculture sustainability. Moreover, very recent studies have demonstrated that, besides improving mineral nutrition, the use of PGPR as plant biostimulants can also positively affect the quality and the nutraceutical values of agricultural products (Kolega et al., 2020; Pii et al., 2018).
Astolfi, S., Pii, Y., Mimmo, T., Lucini, L., Miras-moreno, M. B., Coppa, E., et al. (2020). Single and Combined Fe and S Deficiency Differentially Modulate Root Exudate Composition in Tomato : A Double Strategy for Fe Acquisition ? Int. J. Mol. Sci., 21(11), 4038. doi:10.3390/ijms21114038.
Brumbarova, T., Bauer, P., and Ivanov, R. (2015). Molecular mechanisms governing Arabidopsis iron uptake. Trends Plant Sci. 20, 124–133. doi:http://dx.doi.org/10.1016/j.tplants.2014.11.004.
Crecchio, C., Mimmo, T., Bulgarelli, D., Pertot, I., Pii, Y., Perazzolli, M., et al. (2018). “Beneficial Soil Microbiome for Sustainable Agriculture Production,” in, ed. E. Lichtfouse (Cham: Springer International Publishing), 443–481. doi:10.1007/978-3-319-94232-2_9.
de Santiago, A., García-López, A. M., Quintero, J. M., Avilés, M., and Delgado, A. (2013). Effect of Trichoderma asperellum strain T34 and glucose addition on iron nutrition in cucumber grown on calcareous soils. Soil Biol. Biochem. 57, 598–605. doi:10.1016/j.soilbio.2012.06.020.
Hinsinger, P., Bengough, A. G., Vetterlein, D., and Young, I. (2009). Rhizosphere: biophysics, biogeochemistry and ecological relevance. Plant Soil 321, 117–152. doi:10.1007/s11104-008-9885-9.
Kobayashi, T., and Nishizawa, N. K. (2012). Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 63, 131–52. doi:10.1146/annurev-arplant-042811-105522.
Kolega, S., Moreno, B. M., Buffagni, V., Lucini, L., Valentinuzzi, F., Maver, M., et al. (2020). Nutraceutical profiles of two hydroponically grown sweet basil cultivars as affected by the composition of the nutrient solution and the inoculation with Azospirillum brasilense. Front. Plant Sci. 11, 1683. doi:10.3389/FPLS.2020.596000.
Marastoni, L., Pii, Y., Maver, M., Valentinuzzi, F., Cesco, S., and Mimmo, T. (2019). Role of Azospirillum brasilense in triggering different Fe chelate reductase enzymes in cucumber plants subjected to both nutrient deficiency and toxicity. Plant Physiol. Biochem. 136, 118–126. doi:10.1016/j.plaphy.2019.01.013.
Marschner, H., and Römheld, V. (1994). Strategies of plants for acquisition of iron. Plant Soil 165, 261–274. doi:10.1007/BF00008069.
Pii, Y., Graf, H., Valentinuzzi, F., Cesco, S., and Mimmo, T. (2018). The effects of plant growth-promoting rhizobacteria (PGPR) on the growth and quality of strawberries. Acta Hortic., 231–238. doi:10.17660/actahortic.2018.1217.29.
Pii, Y., Marastoni, L., Springeth, C., Fontanella, M. C., Beone, G. M., Cesco, S., et al. (2016). Modulation of Fe acquisition process by Azospirillum brasilense in cucumber plants. Environ. Exp. Bot. 130, 216–225. doi:10.1016/j.envexpbot.2016.06.011.
Pii, Y., Mimmo, T., Tomasi, N., Terzano, R., Cesco, S., and Crecchio, C. (2015a). Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 51, 403–415. doi:10.1007/s00374-015-0996-1.
Pii, Y., Penn, A., Terzano, R., Crecchio, C., Mimmo, T., and Cesco, S. (2015b). Plant-microorganism-soil interactions influence the Fe availability in the rhizosphere of cucumber plants. Plant Physiol. Biochem. 87, 45–52. doi:10.1016/j.plaphy.2014.12.014.
Vives-Peris, V., de Ollas, C., Gómez-Cadenas, A., and Pérez-Clemente, R. M. (2020). Root exudates: from plant to rhizosphere and beyond. Plant Cell Rep. 39, 3–17. doi:10.1007/s00299-019-02447-5.
Zhang, H., Sun, Y., Xie, X., Kim, M.-S., Dowd, S. E., and Paré, P. W. (2009). A soil bacterium regulates plant acquisition of iron via deficiency-inducible mechanisms. Plant J. 58, 568–577. doi:10.1111/j.1365-313X.2009.03803.x.
Zhao, L., Wang, F., Zhang, Y., and Zhang, J. (2014). Involvement of Trichoderma asperellum strain T6 in regulating iron acquisition in plants. J. Basic Microbiol. 54, S115–S124. doi:10.1002/jobm.201400148.


©2020 BIOSTIMULANT.COM
ALL RIGHTS RESERVED