
Gli idrolizzati proteici animali e vegetali (A-PHs e V-PHs) ottenuti attraverso l'idrolisi incompleta di biomasse residue e/o rifiuti animali o vegetali sono nuove categorie di prodotti dell'economia circolare per l'agricoltura in grado di incrementare in modo sicuro e sostenibile la qualità e la resa degli alimenti (Calvo et al., 2014). I PH sono costituiti principalmente da polipeptidi, oligopeptidi e aminoacidi (Calvo et al., 2014; Colla et al., 2015), carboidrati, fenoli, fitormoni, altri composti organici e minerali in tracce (Colla et al., 2015; Ertani et al., 2014). I loro processi di produzione richiedono un'idrolisi parziale incompleta, termica, enzimatica o chimica (idrolisi alcalina o acida) di rifiuti animali ricchi di proteine e di fonti di biomassa vegetale (Calvo et al., 2014; Colla et al., 2015; du Jardin, 2015). La composizione dei PH varia a seconda della fonte animale o vegetale delle proteine e/o dei processi di produzione (Colla et al., 2017b; Ertani et al., 2009; Ertani et al., 2013). Una volta ottenuti, possono essere utilizzati per via radicale, per applicazione fogliare e per il priming dei semi (Colla et al., 2015; Sorrentino et al., 2021).
Gli A-PH derivano per lo più da sottoprodotti di macellazione, attraverso l'idrolisi chimica del collagene ad alte temperature (ad esempio, 100 °C). Nonostante le preoccupazioni etiche e di sicurezza alimentare, attualmente rappresentano circa il 90% dei PH disponibili sul mercato per la loro comprovata capacità di aumentare l'efficienza d'uso dell'azoto (NUE), la crescita e le difese naturali delle piante, migliorando la tolleranza allo stress salino, allo stress idrico, alle temperature estreme e ai patogeni. Una delle ragioni della loro capacità di esercitare effetti protettivi contro lo stress può essere legata alla loro elevata concentrazione di aminoacidi termostabili come idrossiprolina, idrossilisina, prolina, alanina e glicina (Rouphael et al., 2021). L'idrossiprolina e l'idrossilisina sono costituenti abbondanti del collagene animale e sono responsabili delle sue proprietà funzionali aumentando la reticolazione (Amirrah et al., 2022). Nelle piante, l'idrossiprolina partecipa alla struttura delle glicoproteine ricche di idrossiprolina (HRGPs), i cui prodotti di accoppiamento ossidativo tra residui fenolici e aminoacidi sono essenziali per il rafforzamento delle pareti cellulari e la resistenza agli agenti patogeni (Deepak et al., 2010). La prolina è invece un osmolita compatibile ubiquitario e uno scavenger di ROS, che svolge un ruolo fondamentale nella stabilizzazione delle macromolecole, delle membrane e del potenziale redox cellulare in condizioni di siccità, sale e/o stress ossidativo. Inoltre, può modulare l'espressione di geni che contengono nei loro promotori elementi di risposta alla prolina o all'ipo-osmolarità (PRE), necessari per l'espressione dell'enzima prolina deidrogenasi (PDH) (Carillo, 2018). Tuttavia, gli A-PH sono benefici e convenienti solo se forniti a basse dosi, mentre dosi elevate o trattamenti fogliari ripetuti possono essere tossici (Liu e Lee, 2012). Infatti, poiché il processo di produzione richiede acidi o alcali, gli A-PH contengono alte concentrazioni di cloruro e altri sali che possono indurre stress osmotico e/o ionico. Inoltre, gli A-PH derivati dall'idrolisi chimica possono contenere una grande quantità di aminoacidi liberi in forma D, considerati di basso valore agronomico. Gli alti contenuti di glicina negli A-PH, se forniti per via radicale, possono aumentare l'attività dell'acido 1-aminociclopropano-1-carbossilico sintasi (ACS) e dell'ossidasi (ACO) e, di conseguenza, il contenuto di etilene, riducendo così l'allungamento delle radici e la RUE (Han et al., 2018) e aumentando la senescenza delle foglie. Anche la prolina, se fornita ad alte concentrazioni (≥ 10 mM), può esercitare effetti negativi come l'inibizione dell'allungamento degli ipocotili in Arabidopsis (Hare et al., 2001), o il rapido aumento del cloruro a concentrazioni tossiche nelle foglie di pomodoro (Hayat et al., 2012). Probabilmente gli effetti negativi causati dalla prolina esogena ad alte dosi possono dipendere dall'inibizione a feedback degli enzimi coinvolti nella sua biosintesi nel citosol. Infatti, la stessa sintesi di prolina dal glutammato è in grado di regolare con precisione diversi processi di sviluppo (ad esempio, la formazione dell'embrione, l'allungamento della radice, il tempo di fioritura e la fertilità del polline) attraverso un meccanismo non noto indipendente dagli ormoni (Trovato et al., 2018). Come per la prolina, anche l'idrossiprolina e la glicina betaina possono causare un'inibizione a feedback della loro stessa sintesi. Invece, elevati contenuti di idrossilisina possono inibire l'espressione delle isoforme GS1 e GS2 della glutammina sintetasi nelle foglie di pisello (Leason et al., 1982) e di mais (Acaster e Weitzman, 1985). Inoltre, uno studio di Bernabei (2015) ha dimostrato che il processo di produzione di A-PHs ha un costo energetico più elevato e un maggiore impatto ambientale (consumo di acqua ed emissione di anidride carbonica) rispetto al processo di produzione di idrolizzati vegetali ottenuti mediante idrolisi enzimatica (Bernabei, 2015). Rouphael et al. (2021) hanno dimostrato che le piante di basilico trattate con dosi elevate di un A-PH commerciale accumulavano alti livelli di Na, Cl e prolina. Data l'impossibilità di sintetizzare ulteriore prolina per far fronte agli alti livelli di ioni tossici, le piante sono state costrette a promuovere vie metaboliche alternative per sintetizzare in particolare GABA, che è in grado di agire come osmolita modulando il potenziale osmotico tra citosol e vacuolo, scavenger di ROS e tampone dell'acidosi citoplasmatica (Rouphael et al., 2021). In effetti, ciò distoglie gli scheletri di carbonio dalla crescita, riducendo la resa delle piante.
Al contrario, i trattamenti fogliari con V-PHs, anche a dosi significativamente più elevate di quelle suggerite dai produttori, non hanno mai causato sintomi di fitotossicità (Cerdán et al., 2009; Colla et al., 2014; Kim et al., 2019). Inoltre, per produrre i V-PH possono essere utilizzati anche residui colturali ricchi di proteine o sottoprodotti dell'industria agroalimentare, che rappresentano una materia prima sostenibile ed economica che, se riciclata, contribuisce a ridurre l'impatto ambientale e i costi di produzione (Colla et al. 2017b; Carillo e Morrone 2017). Inoltre, il riutilizzo dei rifiuti biogeni di origine vegetale, evitando il loro inutile smaltimento in discarica, è fondamentale per raggiungere gli obiettivi di economia circolare del Green Deal dell'UE (Commissione Europea, 2019). L'uso della proteolisi enzimatica per ottenere i V-PH permette di applicare parametri di lavorazione come valori di pH neutri e temperature inferiori a 60 °C che evitano la decomposizione degli aminoacidi termolabili (ad esempio, gli aminoacidi primari asparagina, aspartato, glutammato, glutammina e gli aminoacidi essenziali arginina, glicina e istidina) (Colla et al., 2015). Questi ultimi aminoacidi, insieme agli oligopeptidi e ai polipeptidi contenuti nei V-PH, si sono dimostrati in grado di promuovere la risposta alla radicazione (Ceccarelli et al., 2021), di aumentare l'efficienza d'uso delle risorse vegetali (RUE) (Colla et al., 2017a; Rouphael et al., 2021; Rouphael e Colla, 2018), di modulare le attività metaboliche dell'azoto e del carbonio (Colla et al, 2015; Ertani et al., 2013), incrementare la resa (Colla et al., 2017a; Rouphael e Colla, 2018; Sestili et al., 2018), migliorare la qualità nutrizionale e nutraceutica (Paul et al., 2019; Rouphael e Colla, 2018) e aumentare la tolleranza agli stress abiotici (Colla et al., 2017b; Ertani et al., 2013; Lucini et al., 2018; Sorrentino et al., 2021). In effetti, questi effetti non possono essere attribuiti all'azoto organico V-PHs fornito a causa delle dosi estremamente basse utilizzate, che non sono paragonabili a quelle contenute nei comuni fertilizzanti azotati (Halpern et al. 2015). Inoltre, è stato dimostrato che gli effetti dei V-PHs sulla crescita delle radici e dei germogli e la modulazione plastica dell'architettura dell'apparato radicale (ad esempio, lunghezza, numero, densità e superficie delle radici laterali) possono essere dovuti ai peptidi bioattivi che i V-PHs contengono, in grado di esercitare attività fitormonali simili all'auxina e/o alla gibberellina e di agire come molecole di segnalazione (Colla et al., 2015; Ertani et al., 2009; Lucini et al., 2018). Infatti, la maggiore capacità di queste radici modificate di esplorare la rizosfera, insieme alla capacità dei peptidi e degli amminoacidi V-PHs di complessare i nutrienti nel suolo e di aumentare l'attività microbica, accelerano la disponibilità dei nutrienti per l'assorbimento radicale, migliorando la RUE della pianta e la capacità di crescita (Carillo et al., 2019a; Colla et al., 2017b). Inoltre, i V-PH, se somministrati per via radicale, possono sostituire le funzioni dei chelanti sintetici del ferro, evitando il rischio della loro elevata mobilità nel profilo del suolo (Cristofano et al., 2021). Il glutammato, presente in alta concentrazione nei V-PH, può agire come chelante organico migliorando l'assorbimento del ferro e promuovendo l'attività della FeIII-chelato reduttasi sia nelle radici che nei germogli, aumentando così la concentrazione di ferro nelle foglie e l'attività fotosintetica soprattutto nelle giovani piantine (Cerdán et al., 2013; Jeong et al., 2008; Olanrewaju et al., 2019). Infatti, è stato dimostrato che l'uso di V-PHs è in grado di migliorare il contenuto di ferro (e in generale di ioni), lo stato cromatico, la fotosintesi, la produzione e la qualità delle foglie di basilico, spinaci e rucola muraiola perenne (Carillo et al., 2019b; Caruso et al., 2019; Rouphael et al., 2021). Tutti questi effetti benefici esercitati dai V-PH potrebbero essere descritti come "risposta di acquisizione dei nutrienti" e si traducono in un miglioramento generale dell'assorbimento, della traslocazione e dell'assimilazione dei nutrienti (Carillo et al., 2019b; Cristofano et al., 2022; Sestili et al., 2018).
È innegabile che entrambi i tipi di biostimolanti siano validi integratori delle pratiche agricole in grado di aumentare la resa delle colture e la qualità dei prodotti. Tuttavia, mentre per i V-PH non ci sono effetti negativi, è necessario prestare molta attenzione al dosaggio degli A-PH per evitare disturbi al metabolismo e alla crescita e ottimizzare la risposta delle colture (Figura 1).
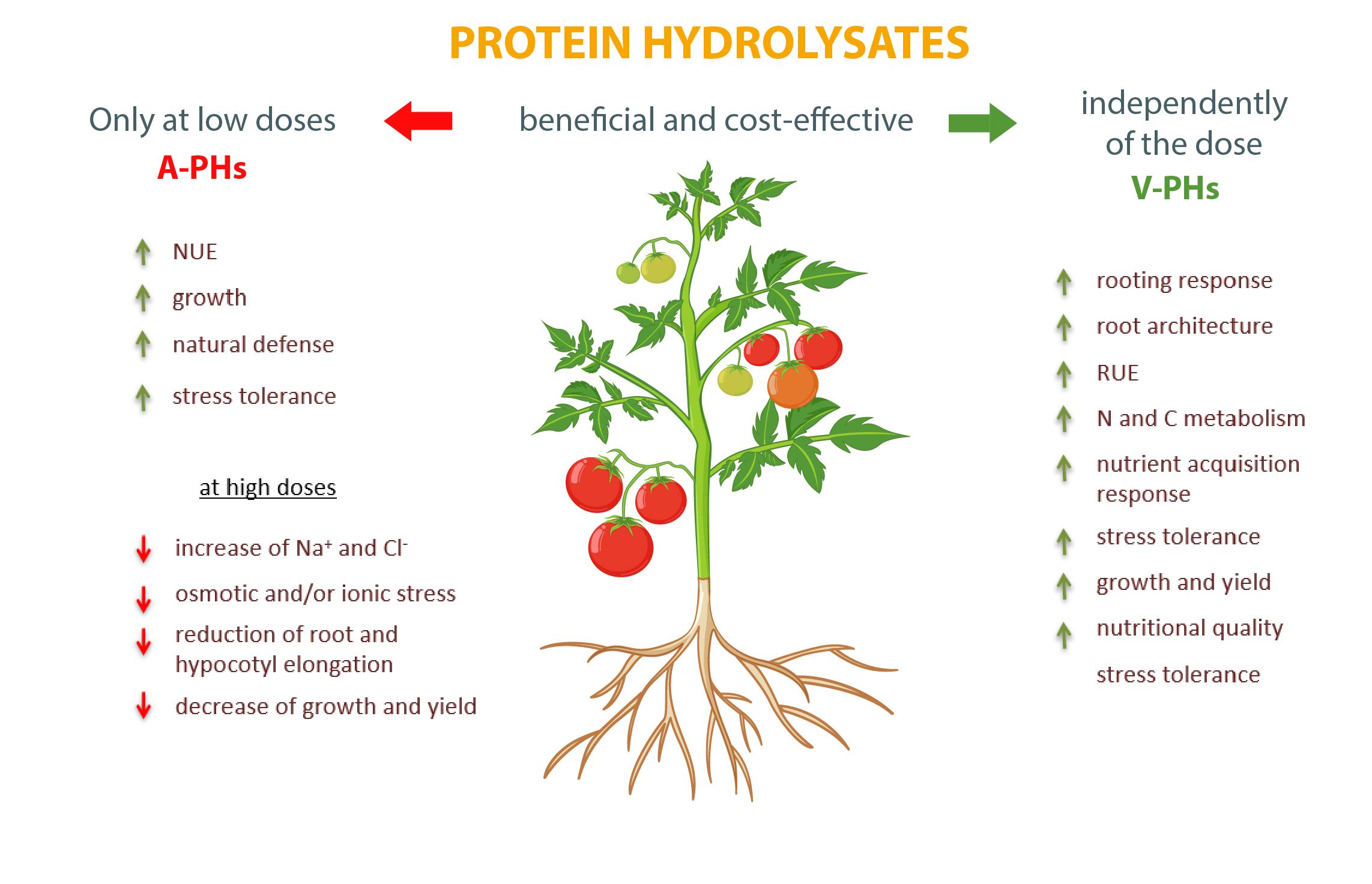
Fig. 1. Principali effetti dose-dipendenti esercitati dagli idrolizzati di proteine animali e vegetali (A-PHs e V-PHs) sulle colture orticole.


© BIOSTIMULANT.COM
TUTTI I DIRITTI RISERVATI