
Les hydrolysats de protéines animales et végétales (A-PHs et V-PHs) obtenus par hydrolyse incomplète de biomasses résiduelles et/ou de déchets animaux ou végétaux sont de nouvelles catégories de produits de l'économie circulaire pour l'agriculture, capables d'augmenter de manière sûre et durable la qualité et le rendement des aliments (Calvo et al., 2014). Les PH sont principalement constitués de polypeptides, d'oligopeptides et d'acides aminés (Calvo et al., 2014 ; Colla et al., 2015), d'hydrates de carbone, de phénols, de phytohormones, d'autres composés organiques et d'oligoéléments (Colla et al., 2015 ; Ertani et al., 2014). Leurs procédés de fabrication nécessitent une hydrolyse partielle incomplète, thermique, enzymatique ou chimique (hydrolyse alcaline ou acide) de déchets animaux riches en protéines et de sources de biomasse végétale (Calvo et al., 2014 ; Colla et al., 2015 ; du Jardin, 2015). La composition des PHs varie en fonction de la source animale ou végétale des protéines et/ou des procédés de fabrication (Colla et al., 2017b ; Ertani et al., 2009 ; Ertani et al., 2013). Une fois obtenus, ils peuvent être utilisés par arrosage des racines, application foliaire et amorçage des semences (Colla et al., 2015 ; Sorrentino et al., 2021).
Les A-PH proviennent principalement de sous-produits d'abattoirs, par hydrolyse chimique du collagène à des températures élevées (par exemple, 100 °C). Malgré les préoccupations éthiques et de sécurité alimentaire, ils représentent actuellement environ 90 % des PH disponibles sur le marché en raison de leur capacité avérée à augmenter l'efficacité de l'utilisation de l'azote (NUE), la croissance des plantes et les défenses naturelles des plantes, en améliorant la tolérance au stress salin, au stress hydrique, aux températures extrêmes et aux agents pathogènes. L'une des raisons de leur capacité à exercer des effets protecteurs contre le stress peut être liée à leur forte concentration en acides aminés thermostables tels que l'hydroxyproline, l'hydroxylysine, la proline, l'alanine et la glycine (Rouphael et al., 2021). L'hydroxyproline et l'hydroxylysine sont des constituants abondants du collagène animal et sont responsables de ses propriétés fonctionnelles en augmentant la réticulation (Amirrah et al., 2022). Chez les plantes, l'hydroxyproline participe à la structure des glycoprotéines riches en hydroxyproline (HRGP), dont les produits de couplage oxydatif des résidus phénoliques et acides aminés sont essentiels pour le renforcement des parois cellulaires et la résistance aux pathogènes (Deepak et al., 2010). En revanche, la proline est un osmolyte compatible omniprésent et un piégeur de ROS, qui joue un rôle essentiel dans la stabilisation des macromolécules, des membranes et du potentiel redox cellulaire en cas de sécheresse, de sel et/ou de stress oxydatif. En outre, il peut moduler l'expression des gènes contenant de la proline ou un élément sensible à l'hypo-osmolarité (PRE) dans leurs promoteurs, nécessaire à l'expression de l'enzyme proline déshydrogénase (PDH) (Carillo, 2018). Cependant, les A-PH ne sont bénéfiques et rentables que lorsqu'ils sont apportés à faible dose, alors que des doses élevées ou des traitements foliaires répétés peuvent être toxiques (Liu et Lee, 2012). En effet, le processus de fabrication nécessitant des acides ou des alcalis, les A-PH contiennent de fortes concentrations de chlorure et d'autres sels qui peuvent induire un stress osmotique et/ou ionique. De plus, les A-PH dérivés de l'hydrolyse chimique peuvent contenir une grande quantité d'acides aminés libres sous forme D qui sont considérés comme ayant une faible valeur agronomique. Les teneurs élevées en glycine dans les A-PH, lorsqu'elles sont fournies par arrosage des racines, peuvent augmenter l'activité de l'acide 1-aminocyclopropane-1-carboxylique synthase (ACS) et de l'oxydase (ACO) et, par conséquent, la teneur en éthylène, réduisant ainsi l'élongation des racines et l'URE (Han et al., 2018) et augmentant la sénescence des feuilles. De même, la proline, si elle est fournie à des concentrations élevées (≥ 10 mM), peut exercer des effets négatifs tels que l'inhibition de l'élongation des hypocotyles chez Arabidopsis (Hare et al., 2001), ou l'augmentation rapide du chlorure à des concentrations toxiques dans les feuilles de tomate (Hayat et al., 2012). Les effets négatifs causés par la proline exogène à des doses élevées peuvent probablement dépendre de l'inhibition en retour des enzymes impliquées dans sa biosynthèse dans le cytosol. En fait, la synthèse de la proline à partir du glutamate lui-même est capable de régler avec précision plusieurs processus de développement (par exemple, la formation de l'embryon, l'élongation des racines, le temps de floraison et la fertilité du pollen) par un mécanisme inconnu et indépendant des hormones (Trovato et al., 2018). Comme pour la proline, l'hydroxyproline et la glycine-bétaïne peuvent également provoquer une inhibition en retour de leur propre synthèse. En revanche, des teneurs élevées en hydroxylysine peuvent inhiber l'expression des isoformes GS1 et GS2 de la glutamine synthétase dans les feuilles de pois (Leason et al., 1982) et de maïs (Acaster et Weitzman, 1985). De plus, une étude de Bernabei (2015), a montré que le procédé de production des A-PHs a un coût énergétique plus élevé et un impact environnemental plus important (consommation d'eau et émission de dioxyde de carbone) que le procédé de production des hydrolysats végétaux obtenus par hydrolyse enzymatique (Bernabei, 2015). Rouphael et al. (2021) ont démontré que des plants de basilic traités avec de fortes doses d'un A-PH commercial accumulaient des niveaux élevés de Na, Cl et proline. Étant donné l'impossibilité de synthétiser davantage de proline pour faire face aux niveaux élevés d'ions toxiques, les plantes ont été contraintes de promouvoir des voies métaboliques alternatives pour synthétiser en particulier le GABA, qui est capable d'agir comme osmolyte modulant le potentiel osmotique entre le cytosol et la vacuole, piégeur de ROS et tampon de l'acidose cytoplasmique (Rouphael et al., 2021). En effet, cela détourne les squelettes de carbone de la croissance, réduisant ainsi le rendement de la plante.
Au contraire, les traitements foliaires de V-PHs, même à des doses significativement plus élevées que celles suggérées par les fabricants, n'ont jamais provoqué de symptômes de phytotoxicité (Cerdán et al., 2009 ; Colla et al., 2014 ; Kim et al., 2019). En outre, pour produire des V-PH, on peut également utiliser des résidus de culture riches en protéines ou des sous-produits de l'industrie agroalimentaire, qui représentent une matière première durable et économique qui, si elle est recyclée, permet de réduire l'impact environnemental et les coûts de production (Colla et al. 2017b ; Carillo et Morrone 2017). En outre, la réutilisation des déchets biogènes d'origine végétale, en évitant leur mise en décharge inutile, est essentielle pour atteindre les objectifs de l'économie circulaire du Green Deal de l'UE (Commission européenne, 2019). L'utilisation de la protéolyse enzymatique pour obtenir des V-PH permet d'appliquer des paramètres de traitement tels que des valeurs de pH neutres et des températures inférieures à 60 °C qui évitent la décomposition des acides aminés thermolabiles (par exemple, les acides aminés primaires asparagine, aspartate, glutamate, glutamine, et les acides aminés essentiels arginine, glycine et histidine) (Colla et al., 2015). Ces derniers acides aminés, ainsi que les oligopeptides et les polypeptides contenus dans les V-PH, se sont révélés capables de favoriser la réaction d'enracinement (Ceccarelli et al., 2021), d'accroître l'efficacité de l'utilisation des ressources végétales (RUE) (Colla et al., 2017a ; Rouphael et al., 2021 ; Rouphael et Colla, 2018), de moduler les activités métaboliques de l'azote et du carbone (Colla et al., 2015 ; Ertani et al., 2013), augmenter le rendement (Colla et al., 2017a ; Rouphael et Colla, 2018 ; Sestili et al., 2018), améliorer la qualité nutritionnelle et nutraceutique (Paul et al., 2019 ; Rouphael et Colla, 2018) et accroître la tolérance au stress abiotique (Colla et al., 2017b ; Ertani et al., 2013 ; Lucini et al., 2018 ; Sorrentino et al., 2021). En effet, ces effets ne peuvent pas être attribués à l'azote organique V-PHs fourni en raison des doses extrêmement faibles utilisées, qui ne sont pas comparables à celles contenues dans les engrais azotés courants (Halpern et al. 2015). En outre, il a été prouvé que les effets des V-PH sur la croissance des racines et des pousses et la modulation plastique de l'architecture du système racinaire (par exemple, la longueur, le nombre, la densité et la surface des racines latérales) peuvent être dus aux peptides bioactifs que contiennent les V-PH, capables d'exercer des activités phytohormonales similaires à l'auxine et/ou à la gibbérelline et d'agir comme des molécules de signalisation (Colla et al., 2015 ; Ertani et al., 2009 ; Lucini et al., 2018). En effet, la plus grande capacité de ces racines modifiées à explorer la rhizosphère, ainsi que la capacité des peptides et des acides aminés V-PHs à complexer les nutriments dans le sol et à augmenter l'activité microbienne, accélèrent la disponibilité des nutriments pour l'absorption par les racines, améliorent l'URE de la plante et sa capacité de croissance (Carillo et al., 2019a ; Colla et al., 2017b). En outre, les V-PH, lorsqu'ils sont fournis par arrosage des racines, peuvent remplacer les fonctions des chélateurs synthétiques du fer en évitant le risque de leur grande mobilité dans le profil du sol (Cristofano et al., 2021). Le glutamate, présent en forte concentration dans les V-PH, peut agir comme un chélateur organique améliorant l'absorption du fer et favorisant l'activité de la FeIII-chelate réductase à la fois dans les racines et les pousses, augmentant ainsi la concentration de fer dans les feuilles et l'activité photosynthétique, en particulier chez les jeunes semis (Cerdán et al., 2013 ; Jeong et al., 2008 ; Olanrewaju et al., 2019). En fait, l'utilisation des V-PH s'est avérée capable d'améliorer la teneur en fer (et en général en ions), l'état de la couleur, la photosynthèse, la production et la qualité des feuilles de basilic, d'épinard et de roquette vivace (Carillo et al., 2019b ; Caruso et al., 2019 ; Rouphael et al., 2021). Tous ces effets bénéfiques exercés par les V-PH pourraient être décrits comme une "réponse à l'acquisition des nutriments" et se traduisent par une amélioration générale de l'absorption, de la translocation et de l'assimilation des nutriments (Carillo et al., 2019b ; Cristofano et al., 2022 ; Sestili et al., 2018).
Il est indéniable que les deux types de biostimulants sont des compléments valables aux pratiques agricoles, capables d'augmenter le rendement des cultures et la qualité des produits. Cependant, alors que les V-PH n'ont pas d'effets indésirables, il est nécessaire de faire très attention au dosage des A-PH afin d'éviter les troubles du métabolisme et de la croissance et d'optimiser la réponse des cultures (figure 1).
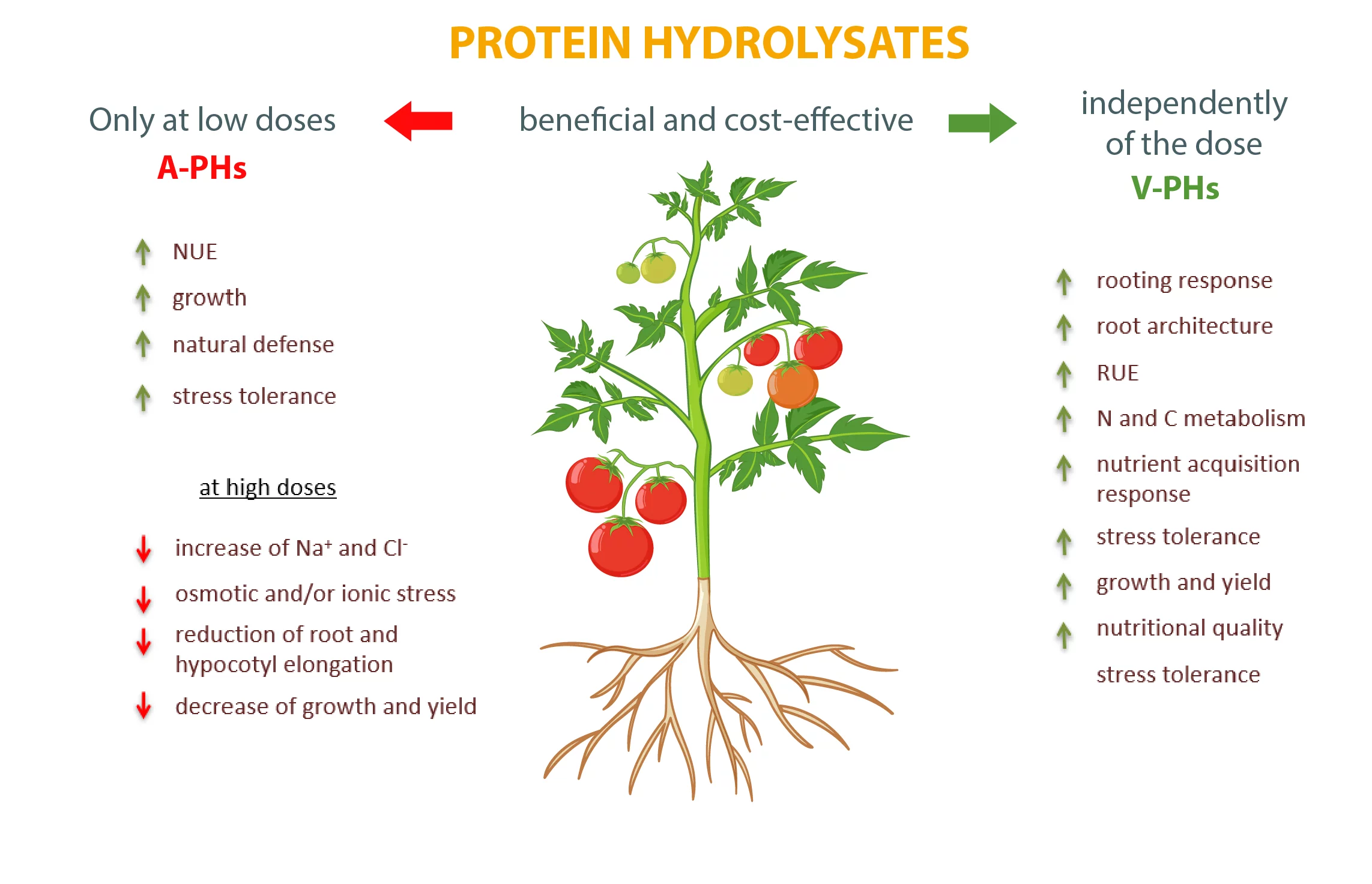
Fig. 1. Principaux effets dose-dépendants exercés par les hydrolysats de protéines animales et végétales (A-PHs et V-PHs) sur les cultures légumières.


© BIOSTIMULANT.COM
TOUS DROITS RÉSERVÉS