
Гидролизаты белков животного и растительного происхождения (A-PHs и V-PHs), полученные в результате неполного гидролиза остаточных биомасс и/или отходов животного или растительного происхождения, являются новыми категориями продукции с циркулярной экономикой для сельского хозяйства, способными безопасно и устойчиво повысить качество и урожайность продуктов питания (Calvo et al., 2014). РН в основном состоят из полипептидов, олигопептидов и аминокислот (Calvo et al., 2014; Colla et al., 2015), углеводов, фенолов, фитогормонов, других органических соединений и микроэлементов (Colla et al., 2015; Ertani et al., 2014). Процессы их производства требуют неполного термического, ферментативного или химического (щелочной или кислотный гидролиз) частичного гидролиза богатых белком отходов животного происхождения и растительной биомассы (Calvo et al., 2014; Colla et al., 2015; du Jardin, 2015). Состав РН варьируется в зависимости от животного или растительного источника белков и/или процессов производства (Colla et al., 2017b; Ertani et al., 2009; Ertani et al., 2013). После получения они могут быть использованы путем корневой подкормки, внекорневого внесения и грунтования семян (Colla et al., 2015; Sorrentino et al., 2021).
А-РН в основном получают из побочных продуктов скотобоен, путем химического гидролиза коллагена при высоких температурах (например, 100 °C). Несмотря на этические соображения и проблемы безопасности пищевых продуктов, в настоящее время они составляют около 90% РН, доступных на рынке, благодаря своей доказанной способности повышать эффективность использования азота (NUE), рост растений и естественную защиту растений, улучшая устойчивость к солевому стрессу, водному стрессу, экстремальным температурам и патогенам. Одна из причин их способности оказывать стрессозащитное действие может быть связана с высокой концентрацией термостабильных аминокислот, таких как гидроксипролин, гидроксилизин, пролин, аланин и глицин (Rouphael et al., 2021). Гидроксипролин и гидроксилизин являются многочисленными компонентами животного коллагена и отвечают за его функциональные свойства, увеличивая поперечные связи (Amirrah et al., 2022). В растениях гидроксипролин участвует в структуре гидроксипролин-богатых гликопротеинов (HRGPs), чьи продукты окислительного соединения остатков фенол-аминокислот необходимы для укрепления клеточных стенок и устойчивости к патогенам (Deepak et al., 2010). Напротив, пролин является вездесущим совместимым осмолитом и поглотителем ROS, который играет ключевую роль в стабилизации макромолекул, мембран и клеточного окислительно-восстановительного потенциала в условиях засухи, солевого и/или окислительного стресса. Кроме того, он может модулировать экспрессию генов, содержащих в своих промоторах пролин или гипоосмолярно-реактивный элемент (PRE), необходимый для экспрессии фермента пролиндегидрогеназы (PDH) (Carillo, 2018). Однако A-PH являются полезными и экономически эффективными только при внесении в низких дозах, в то время как высокие дозы или повторные внекорневые обработки могут быть токсичными (Liu and Lee, 2012). Фактически, поскольку процесс производства требует использования кислот или щелочей, A-PH содержат высокие концентрации хлоридов и других солей, которые могут вызывать осмотический и/или ионный стресс. Более того, A-PH, полученные в результате химического гидролиза, могут содержать большое количество свободных аминокислот в D-форме, которые считаются малоценными в агрономическом отношении. Высокое содержание глицина в A-PH, когда он поступает с корневой заправкой, может увеличить активность 1-аминоциклопропан-1-карбоновой кислоты синтазы (ACS) и оксидазы (ACO) и, следовательно, содержание этилена, тем самым снижая удлинение корней и RUE (Han et al., 2018) и усиливая старение листьев. Также пролин, если его поставлять в высоких концентрациях (≥ 10 мМ), может оказывать негативные эффекты, такие как ингибирование удлинения гипокотилей у арабидопсиса (Hare et al., 2001) или быстрое увеличение содержания хлоридов в токсичных концентрациях в листьях томата (Hayat et al., 2012). Вероятно, негативные эффекты, вызываемые экзогенным пролином в высоких дозах, могут зависеть от обратного ингибирования ферментов, участвующих в его биосинтезе в цитозоле. На самом деле, синтез пролина из глутамата способен регулировать некоторые процессы развития (например, формирование зародыша, удлинение корня, время цветения и фертильность пыльцы) с помощью неизвестного гормон-независимого механизма (Trovato et al., 2018). Как и пролин, гидроксипролин и глицин бетаин могут вызывать ингибирование собственного синтеза по обратной связи. В то время как высокое содержание гидроксилизина может ингибировать экспрессию изоформ GS1 и GS2 глутаминсинтетазы в листьях мочи (Leason et al., 1982) и кукурузы (Acaster and Weitzman, 1985). Кроме того, исследование Bernabei (2015), показало, что процесс производства A-PHs имеет более высокие энергетические затраты и большее воздействие на окружающую среду (потребление воды и выброс углекислого газа), чем процесс производства растительных гидролизатов, полученных ферментативным гидролизом (Bernabei, 2015). Rouphael et al. (2021) продемонстрировали, что растения базилика, обработанные высокими дозами коммерческого A-PH, накапливали высокие уровни Na, Cl и пролина. Поскольку невозможно было синтезировать пролин, чтобы справиться с высоким уровнем токсичных ионов, растения были вынуждены использовать альтернативные метаболические пути для синтеза, в частности, ГАМК, которая может действовать как осмолит, модулирующий осмотический потенциал между цитозолем и вакуолями, поглотитель ROS и буфер цитоплазматического ацидоза (Rouphael et al., 2021). Действительно, это отвлекает углеродные скелеты от роста, снижая урожайность растений.
Напротив, внекорневые обработки V-PHs, в том числе в значительно более высоких дозах, чем предлагаемые производителями, никогда не вызывали симптомов фитотоксичности (Cerdán et al., 2009; Colla et al., 2014; Kim et al., 2019). Кроме того, для производства V-PHs могут также использоваться богатые белком растительные остатки или побочные продукты агропищевой промышленности, которые представляют собой устойчивое и экономичное сырье, которое при повторном использовании помогает снизить воздействие на окружающую среду и производственные затраты (Colla et al. 2017b; Carillo and Morrone 2017). Более того, повторное использование биогенных отходов растительного происхождения во избежание их ненужного захоронения имеет ключевое значение для достижения целей "Зеленого курса" ЕС в области циркулярной экономики (Европейская комиссия, 2019). Использование ферментативного протеолиза для получения V-PHs позволяет применять такие параметры обработки, как нейтральные значения pH и температура ниже 60 °C, что позволяет избежать разложения термолабильных аминокислот (например, первичных аминокислот аспарагина, аспартата, глутамата, глутамина и незаменимых аминокислот аргинина, глицина и гистидина) (Colla et al., 2015). Последние аминокислоты, а также олигопептиды и полипептиды, содержащиеся в V-PHs, как было доказано, способны стимулировать реакцию укоренения (Ceccarelli et al., 2021), повышать эффективность использования растительных ресурсов (RUE) (Colla et al., 2017a; Rouphael et al., 2021; Rouphael and Colla, 2018), модулировать метаболизм азота и углерода (Colla et al., 2015; Ertani et al., 2013), повышают урожайность (Colla et al., 2017a; Rouphael and Colla, 2018; Sestili et al., 2018), улучшают качество питательных веществ и нутрицевтиков (Paul et al., 2019; Rouphael and Colla, 2018) и повышают устойчивость к абиотическим стрессам (Colla et al., 2017b; Ertani et al., 2013; Lucini et al., 2018; Sorrentino et al., 2021). Действительно, эти эффекты не могут быть приписаны поставляемому V-PHs органическому азоту из-за чрезвычайно низких доз, которые не сравнимы с теми, что содержатся в обычных азотных удобрениях (Halpern et al. 2015). Кроме того, было доказано, что влияние V-PHs на рост корней и побегов и пластическая модуляция архитектуры корневой системы (например, длина, количество, плотность и поверхность боковых корней) могут быть обусловлены содержащимися в V-PHs биологически активными пептидами, способными оказывать ауксиноподобную и/или гиббереллиноподобную фитогормональную активность и действовать как сигнальные молекулы (Colla et al., 2015; Ertani et al., 2009; Lucini et al., 2018). Действительно, более высокая способность этих модифицированных корней исследовать ризосферу, а также способность пептидов и аминокислот V-PHs комплексовать питательные вещества в почве и повышать микробную активность ускоряют доступность питательных веществ для поглощения корнями, повышают РУЭ и способность растений к росту (Carillo et al., 2019a; Colla et al., 2017b). Кроме того, V-PHs, поставляемые при корневой подкормке, могут заменить функции синтетических хелаторов железа, избегая риска их высокой мобильности в почвенном профиле (Cristofano et al., 2021). Глутамат, присутствующий в высокой концентрации в V-PHs, может действовать как органический хелатор, улучшая поглощение железа и стимулируя активность FeIII-хелат-редуктазы как в корнях, так и в побегах, тем самым увеличивая концентрацию железа в листьях и фотосинтетическую активность, особенно в молодых проростках (Cerdán et al., 2013; Jeong et al., 2008; Olanrewaju et al., 2019). Фактически, было показано, что использование V-PHs способно улучшить содержание железа (и в целом ионов), цветовой статус, фотосинтез, производство и качество в листьях базилика, шпината и многолетней настенной ракеты (Carillo et al., 2019b; Caruso et al., 2019; Rouphael et al., 2021). Все эти полезные эффекты, оказываемые V-PHs, могут быть описаны как "реакция приобретения питательных веществ" и приводят к общему улучшению поглощения, транслокации и ассимиляции питательных веществ (Carillo et al., 2019b; Cristofano et al., 2022; Sestili et al., 2018).
Бесспорно, оба типа биостимуляторов являются эффективными дополнениями к сельскохозяйственной практике, способными повысить урожайность и качество продукции. Однако, в то время как для V-PHs не существует негативных эффектов, необходимо уделять пристальное внимание дозировке A-PHs, чтобы избежать негативных нарушений метаболизма и роста и оптимизировать реакцию культур (Рисунок 1).
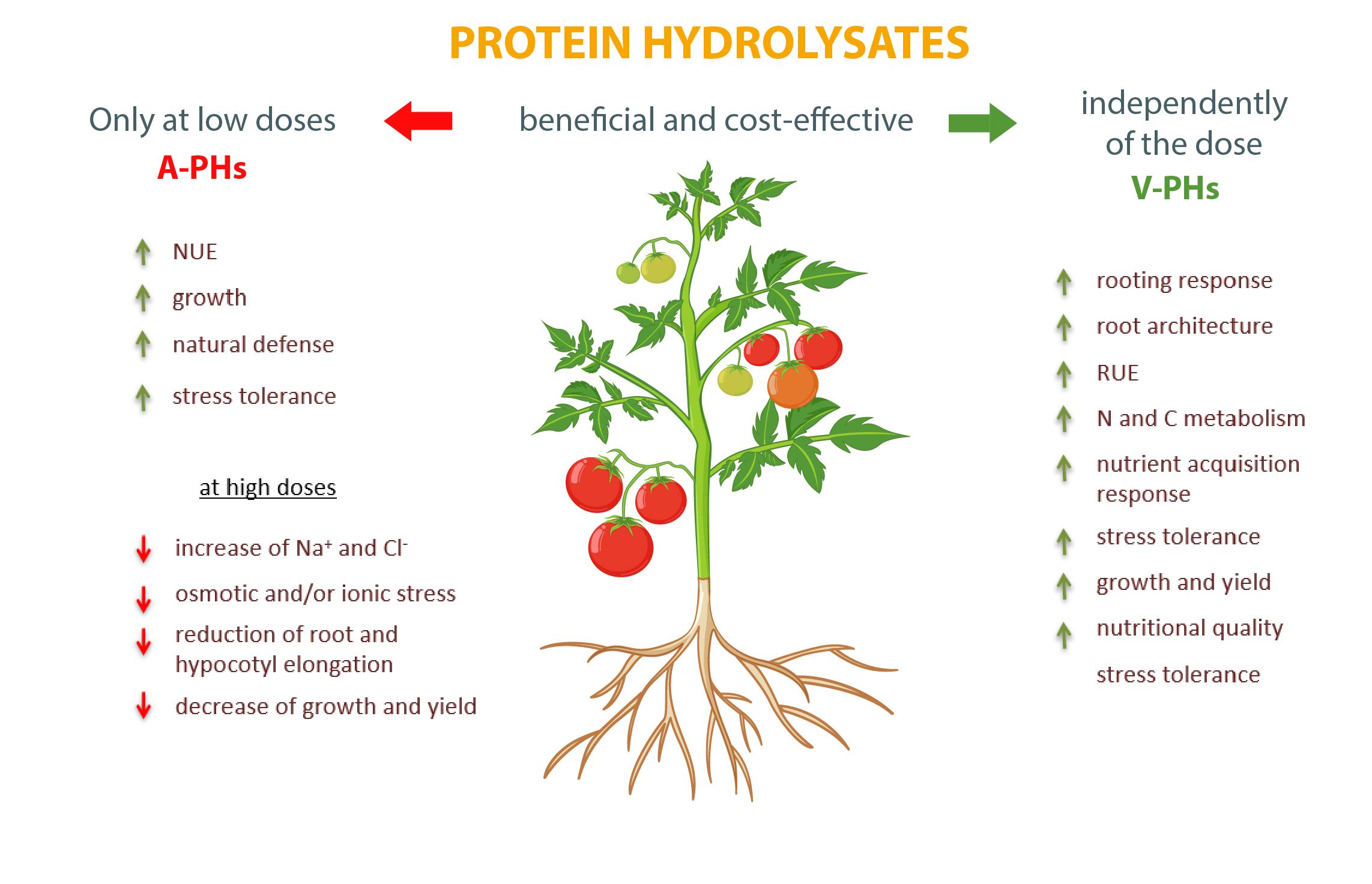
Рис. 1. Основные дозозависимые эффекты, оказываемые гидролизатами животного и растительного белка (A-PHs и V-PHs) на овощные культуры.


©2020 BIOSTIMULANT.COM
ВСЕ ПРАВА ЗАЩИЩЕНЫ