
Los hidrolizados de proteínas animales y vegetales (A-PH y V-PH) obtenidos mediante la hidrólisis incompleta de biomasas residuales y/o residuos animales o vegetales son nuevas categorías de productos de economía circular para la agricultura capaces de aumentar de forma segura y sostenible la calidad y el rendimiento de los alimentos (Calvo et al., 2014). Los PHs están constituidos principalmente por polipéptidos, oligopéptidos y aminoácidos (Calvo et al., 2014; Colla et al., 2015), carbohidratos, fenoles, fitohormonas, otros compuestos orgánicos y minerales traza (Colla et al., 2015; Ertani et al., 2014). Sus procesos de fabricación requieren una hidrólisis parcial incompleta térmica, enzimática o química (hidrólisis alcalina o ácida) de residuos animales ricos en proteínas y fuentes de biomasa vegetal (Calvo et al., 2014; Colla et al., 2015; du Jardin, 2015). La composición de los PHs varía en función de la fuente animal o vegetal de proteínas y/o de los procesos de fabricación (Colla et al., 2017b; Ertani et al., 2009; Ertani et al., 2013). Una vez obtenidos, pueden utilizarse mediante empapado radicular, aplicación foliar e imprimación de semillas (Colla et al., 2015; Sorrentino et al., 2021).
Los A-PH proceden principalmente de subproductos de matadero, mediante hidrólisis química del colágeno a altas temperaturas (por ejemplo, 100 °C). A pesar de las preocupaciones éticas y de seguridad alimentaria, actualmente representan alrededor del 90% de los PHs disponibles en el mercado por su capacidad demostrada para aumentar la eficiencia de uso del nitrógeno (NUE), el crecimiento de las plantas y las defensas naturales de las plantas, mejorando la tolerancia al estrés salino, al estrés hídrico, a las temperaturas extremas y a los patógenos. Una de las razones de su capacidad para ejercer efectos protectores frente al estrés puede estar relacionada con su alta concentración de aminoácidos termoestables como la hidroxiprolina, la hidroxilisina, la prolina, la alanina y la glicina (Rouphael et al., 2021). La hidroxiprolina y la hidroxilisina son constituyentes abundantes del colágeno animal siendo responsables de sus propiedades funcionales al aumentar la reticulación (Amirrah et al., 2022). En las plantas, la hidroxiprolina forma parte de la estructura de las glicoproteínas ricas en hidroxiprolina (HRGPs), cuyos productos de acoplamiento oxidativo de residuos fenólicos-aminoácidos son esenciales para el fortalecimiento de las paredes celulares y la resistencia frente a patógenos (Deepak et al., 2010). En cambio, la prolina es un osmolito compatible ubicuo y un eliminador de ROS, que desempeña un papel fundamental en la estabilización de macromoléculas, membranas y potencial redox celular en condiciones de sequía, sal y/o estrés oxidativo. Además, puede modular la expresión de genes que contienen en sus promotores la prolina o el elemento de respuesta a la hipoosmolaridad (PRE), necesarios para la expresión de la enzima prolina deshidrogenasa (PDH) (Carillo, 2018). Sin embargo, los A-PH son beneficiosos y rentables solo cuando se suministran a dosis bajas, mientras que dosis altas o tratamientos foliares repetidos pueden ser tóxicos (Liu y Lee, 2012). De hecho, dado que el proceso de fabricación requiere ácidos o álcalis, los A-PHs contienen altas concentraciones de cloruro y otras sales que pueden inducir estrés osmótico y/o iónico. Además, los A-PH derivados de la hidrólisis química pueden contener una gran cantidad de aminoácidos libres en forma D que se consideran de bajo valor agronómico. Los altos contenidos de glicina en A-PHs, cuando se suministran por empapamiento radicular, pueden aumentar la actividad de la 1-aminociclopropano-1-ácido carboxílico sintasa (ACS) y oxidasa (ACO) y, por lo tanto, el contenido de etileno, reduciendo así la elongación radicular y la RUE (Han et al., 2018) y aumentando la senescencia foliar. También la prolina, si se suministra a altas concentraciones (≥ 10 mM), puede ejercer efectos negativos como la inhibición de la elongación de los hipocótilos en Arabidopsis (Hare et al., 2001), o el rápido aumento de cloruro a concentraciones tóxicas en hojas de tomate (Hayat et al., 2012). Probablemente, los efectos negativos causados por la prolina exógena a altas dosis pueden depender de la inhibición por retroalimentación de las enzimas implicadas en su biosíntesis en el citosol. De hecho, la propia síntesis de prolina a partir de glutamato es capaz de afinar varios procesos de desarrollo (por ejemplo, la formación de embriones, el alargamiento de las raíces, el tiempo de floración y la fertilidad del polen) por un mecanismo desconocido independiente de las hormonas (Trovato et al., 2018). En cuanto a la prolina, también la hidroxiprolina y la glicina betaína pueden causar inhibición por retroalimentación de su propia síntesis. Mientras que, altos contenidos de hidroxilisina, pueden inhibir la expresión de las isoformas GS1 y GS2 de la glutamina sintetasa en hojas de guisante (Leason et al., 1982) y maíz (Acaster y Weitzman, 1985). Además, un estudio de Bernabei (2015), demostró que el proceso de producción de A-PHs tiene un mayor coste energético y un mayor impacto ambiental (consumo de agua y emisión de dióxido de carbono) que el proceso de producción de hidrolizados vegetales obtenidos por hidrólisis enzimática (Bernabei, 2015). Rouphael et al. (2021) demostraron que las plantas de albahaca tratadas con altas dosis de un A-PH comercial acumulaban altos niveles de Na, Cl y prolina. Ante la imposibilidad de sintetizar más prolina para hacer frente a los altos niveles de iones tóxicos, las plantas se vieron obligadas a promover vías metabólicas alternativas para sintetizar en particular GABA, que es capaz de actuar como osmolito modulador del potencial osmótico entre el citosol y la vacuola, eliminador de ROS y tampón de la acidosis citoplasmática (Rouphael et al., 2021). De hecho, esto desvía los esqueletos de carbono del crecimiento reduciendo el rendimiento de la planta.
Por el contrario, los tratamientos foliares con V-PHs, también a dosis significativamente superiores a las sugeridas por los fabricantes, nunca han causado síntomas de fitotoxicidad (Cerdán et al., 2009; Colla et al., 2014; Kim et al., 2019). Además, para producir V-PHs también se pueden utilizar residuos de cultivos ricos en proteínas o subproductos de la industria agroalimentaria, que representan una materia prima sostenible y económica que, si se recicla, ayuda a reducir el impacto ambiental y los costes de producción (Colla et al. 2017b; Carillo y Morrone 2017). Además, la reutilización de residuos biogénicos derivados de plantas evitando su vertido innecesario es fundamental para cumplir los objetivos de economía circular del Green Deal de la UE (Comisión Europea, 2019). El uso de la proteólisis enzimática para obtener V-PH permite aplicar parámetros de procesamiento como valores de pH neutro y temperaturas inferiores a 60 °C que evitan la descomposición de aminoácidos termolábiles (por ejemplo, los aminoácidos primarios asparagina, aspartato, glutamato, glutamina, y los aminoácidos esenciales arginina, glicina e histidina) (Colla et al., 2015). Estos últimos aminoácidos junto con los oligopéptidos y polipéptidos contenidos en las V-PH, han demostrado ser capaces de promover la respuesta de enraizamiento (Ceccarelli et al., 2021), aumentar la eficiencia de uso de los recursos de la planta (RUE) (Colla et al., 2017a; Rouphael et al., 2021; Rouphael y Colla, 2018), modular las actividades metabólicas de nitrógeno y carbono (Colla et al., 2015; Ertani et al., 2013), impulsar el rendimiento (Colla et al., 2017a; Rouphael y Colla, 2018; Sestili et al., 2018), mejorar la calidad nutricional y nutracéutica (Paul et al., 2019; Rouphael y Colla, 2018) y aumentar la tolerancia al estrés abiótico (Colla et al., 2017b; Ertani et al., 2013; Lucini et al., 2018; Sorrentino et al., 2021). De hecho, estos efectos no pueden atribuirse al nitrógeno orgánico V-PHs suministrado debido a las dosis extremadamente bajas utilizadas, que no son comparables con las contenidas en los fertilizantes nitrogenados comunes (Halpern et al. 2015). Además, se ha comprobado que los efectos de los V-PHs sobre el crecimiento de raíces y brotes y la modulación plástica de la arquitectura del sistema radicular (por ejemplo, longitud, número, densidad y superficie de las raíces laterales) pueden deberse a los péptidos bioactivos que contienen los V-PHs, capaces de ejercer actividades fitohormonales similares a la auxina y/o la giberelina y actuar como moléculas de señalización (Colla et al., 2015; Ertani et al., 2009; Lucini et al., 2018). De hecho, la mayor capacidad de estas raíces modificadas para explorar la rizosfera, junto con la capacidad de los péptidos y aminoácidos V-PHs para complejizar los nutrientes en el suelo y aumentar la actividad microbiana aceleran la disponibilidad de nutrientes para la absorción radicular, mejoran la RUE de la planta y su capacidad de crecimiento (Carillo et al., 2019a; Colla et al., 2017b). Además, los V-PHs, cuando se suministran por empapamiento radicular, pueden sustituir las funciones de los quelantes sintéticos del hierro evitando el riesgo de su alta movilidad en el perfil del suelo (Cristofano et al., 2021). El glutamato, presente en altas concentraciones en los V-PHs, puede actuar como quelante orgánico mejorando la absorción de hierro y promoviendo la actividad FeIII-quelato reductasa tanto en raíces como en brotes, incrementando así la concentración de hierro en hojas y la actividad fotosintética especialmente en plántulas jóvenes (Cerdán et al., 2013; Jeong et al., 2008; Olanrewaju et al., 2019). De hecho, se ha demostrado que el uso de V-PHs puede mejorar el contenido de hierro (y en general de iones), el estado de color, la fotosíntesis, la producción y la calidad en hojas de albahaca, espinaca y rúcula de pared perenne (Carillo et al., 2019b; Caruso et al., 2019; Rouphael et al., 2021). Todos estos efectos beneficiosos ejercidos por las V-PH podrían describirse como "respuesta de adquisición de nutrientes" y se traducen en una mejora general de la captación, translocación y asimilación de nutrientes (Carillo et al., 2019b; Cristofano et al., 2022; Sestili et al., 2018).
Es innegable que ambos tipos de bioestimulantes son complementos válidos de las prácticas agrícolas capaces de potenciar el rendimiento de los cultivos y la calidad de los productos. Sin embargo, mientras que en el caso de los V-PH no existen efectos adversos, es necesario prestar mucha atención a la dosificación de los A-PH para evitar trastornos negativos en el metabolismo y el crecimiento y optimizar la respuesta de los cultivos (Figura 1).
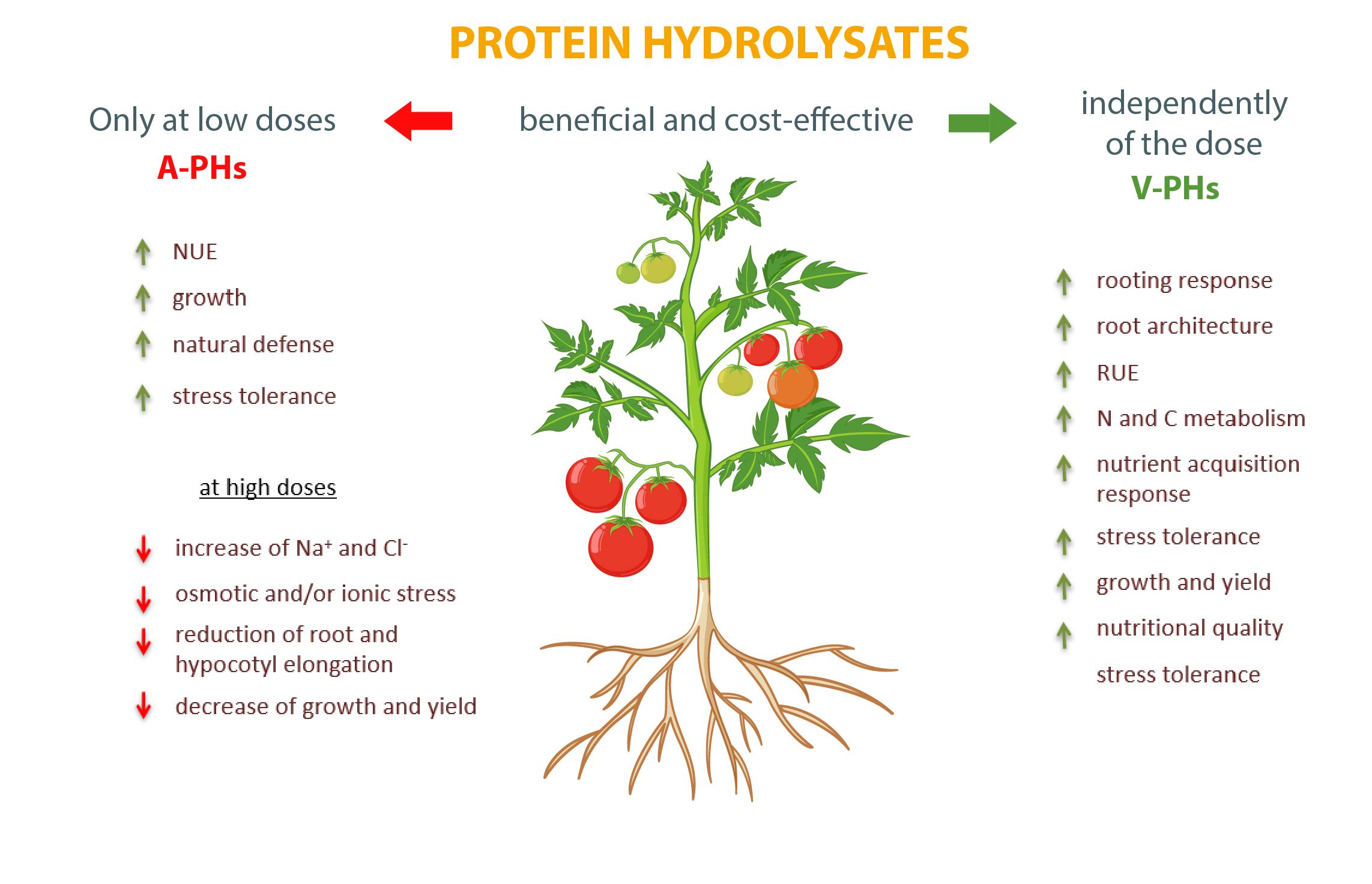
Fig. 1. Principales efectos dosis-dependientes ejercidos por los hidrolizados de proteínas animales y vegetales (A-PHs y V-PHs) sobre los cultivos vegetales.


©2020 BIOSTIMULANT.COM
TODOS LOS DERECHOS RESERVADOS