
通过不完全水解动物或植物残余生物质和/或废物获得的动物和植物蛋白水解物(A-PHs和V-PHs)是新的农业循环经济产品类别,能够安全和可持续地提高食品质量和产量(Calvo等人,2014)。PHs主要由多肽、寡肽和氨基酸(Calvo等人,2014;Colla等人,2015)、碳水化合物、酚类、植物激素、其他有机化合物和微量元素构成(Colla等人,2015;Ertani等人,2014)。它们的制造过程需要对富含蛋白质的动物废弃物和植物生物质来源进行不完全的热、酶或化学(碱性或酸性水解)部分水解(Calvo等人,2014;Colla等人,2015;du Jardin,2015)。PHs的组成根据蛋白质的动物或植物来源和/或制造工艺而变化(Colla等人,2017b;Ertani等人,2009;Ertani等人,2013)。一旦获得,它们可以通过根部浇灌、叶面喷洒和种子打底使用(Colla等人,2015;Sorrentino等人,2021)。
A-PHs大多来自屠宰场的副产品,通过在高温下(如100℃)对胶原蛋白进行化学水解而得来。尽管存在道德和食品安全问题,但目前市场上约有90%的PHs被证明有能力提高氮利用效率(NUE)、植物生长和植物天然防御能力,提高对盐胁迫、水胁迫、极端温度和病原体的耐受性。它们能够发挥压力保护作用的原因之一可能与它们高浓度的恒温氨基酸有关,如羟脯氨酸、羟赖氨酸、脯氨酸、丙氨酸和甘氨酸(Rouphael等,2021)。羟脯氨酸和羟赖氨酸是动物胶原蛋白的丰富成分,通过增加交联负责其功能特性(Amirrah等人,2022)。在植物中,羟脯氨酸参与富含羟脯氨酸的糖蛋白(HRGPs)的结构,其氧化酚-氨基酸残基耦合产物对加强细胞壁和抵抗病原体至关重要(Deepak等人,2010)。相反,脯氨酸是一种无处不在的兼容渗透剂和ROS清除剂,在干旱、盐和/或氧化压力下稳定大分子、膜和细胞氧化还原电位方面起着关键的作用。此外,它可能调节启动子中含有脯氨酸或低渗透性反应元件(PRE)的基因的表达,这是脯氨酸脱氢酶(PDH)的表达所必需的(Carillo, 2018)。然而,A-PHs只有在低剂量提供时才是有益的和具有成本效益的,而高剂量或重复叶面处理可能是有毒的(刘和李,2012)。事实上,由于制造过程需要酸或碱,A-PHs含有高浓度的氯化物和其他盐类,可能诱发渗透和/或离子压力。此外,通过化学水解得到的A-PH可能含有大量的D型游离氨基酸,这些氨基酸被认为具有低农艺价值。A-PH中甘氨酸的高含量,当通过根部浇灌供应时,可能会增加1-氨基环丙烷-1-羧酸合成酶(ACS)和氧化酶(ACO)的活性,从而增加乙烯的含量,从而减少根系伸长和RUE(Han等人,2018)并增强叶片衰老。另外,脯氨酸如果以高浓度(≥10mM)供应,可能会产生负面影响,如抑制拟南芥下胚轴的伸长(Hare等人,2001年),或在番茄叶片中以有毒浓度快速增加氯化物(Hayat等人,2012年)。高剂量的外源脯氨酸引起的负面影响可能取决于参与其在细胞膜上生物合成的酶的反馈抑制。事实上,谷氨酸本身合成的脯氨酸能够通过一种未知的独立于激素的机制微调几个发育过程(如胚胎形成、根系伸长、开花时间和花粉生育能力)(Trovato等人,2018)。至于脯氨酸,还有羟脯氨酸和甘氨酸甜菜碱也可能导致对其自身合成的反馈抑制。 而羟基赖氨酸的高含量,可能会抑制小鼠(Leason等人,1982年)和玉米叶(Acaster和Weitzman,1985年)中谷氨酰胺合成酶GS1和GS2的异构体表达。此外,Bernabei(2015)的一项研究表明,与通过酶水解获得的植物水解物的生产过程相比,A-PHs的生产过程具有更高的能源成本和更大的环境影响(水消耗和二氧化碳排放)(Bernabei,2015)。Rouphael等人(2021)证明,用高剂量的商业A-PH处理的罗勒植物积累了高水平的Na、Cl和脯氨酸。由于不可能进一步合成脯氨酸以应对高水平的有毒离子,植物被迫促进替代代谢途径,以合成特别是GABA,它能够作为渗透压调节细胞膜和液泡之间的渗透势,ROS清除剂和细胞质酸中毒缓冲器(Rouphael等人,2021)。事实上,这转移了碳骨架的生长,降低了植物产量。
相反,V-PHs的叶面处理,也比制造商建议的剂量高得多,但从未引起植物毒性症状(Cerdán等人,2009;Colla等人,2014;Kim等人,2019)。此外,为了生产V-PHs,还可以使用富含蛋白质的作物残体或农业食品工业副产品,这代表了一种可持续的、经济的原材料,如果回收利用,有助于减少环境影响和生产成本(Colla等人,2017b;Carillo和Morrone,2017)。此外,重新利用植物衍生的生物源性废物,避免其不必要的填埋,对于实现欧盟绿色协议的循环经济目标至关重要(欧盟委员会,2019年)。使用酶解法获得V-PHs,可以应用中性pH值和低于60℃的温度等加工参数,避免热分解的氨基酸(如初级氨基酸天冬酰胺、天门冬氨酸、谷氨酸、谷氨酰胺,以及必需氨基酸精氨酸、甘氨酸和组氨酸)的分解(Colla等人,2015)。后面的氨基酸连同V-PHs中包含的寡肽和多肽,已被证明能够促进生根反应(Ceccarelli等人,2021),提高植物资源利用效率(RUE)(Colla等人,2017a;Rouphael等人,2021;Rouphael和Colla,2018),调节氮和碳代谢活动(Colla等人、2015;Ertani等人,2013),提高产量(Colla等人,2017a;Rouphael和Colla,2018;Sestili等人,2018),改善营养和保健品质量(Paul等人,2019;Rouphael和Colla,2018),增加非生物压力耐受性(Colla等人, 2017b;Ertani等人,2013;Lucini等人,2018;Sorrentino等人,2021)。事实上,这些影响不能归因于所提供的V-PHs有机氮,因为所使用的剂量极低,无法与普通氮肥所含的剂量相比(Halpern等人,2015)。此外,已经证明V-PHs对根和芽生长的影响以及对根系结构(如侧根的长度、数量、密度和表面)的塑性调节,可能是由于V-PHs含有的生物活性肽,能够发挥类似辅助素和/或赤霉素的植物激素活性,并作为信号分子发挥作用(Colla等人,2015;Ertani等人,2009;Lucini等人,2018)。事实上,这些修饰过的根有更高的探索能力,加上V-PHs肽和氨基酸在土壤中复合养分的能力,增加微生物的活性加快了根部吸收养分的可用性,提高了植物的RUE和生长能力(Carillo等人,2019a;Colla等人,2017b)。此外,V-PHs通过根部浇灌供应时,可以取代合成铁螯合剂的功能,避免其在土壤剖面中的高流动性风险(Cristofano等人,2021)。在V-PHs中存在高浓度的谷氨酸,可以作为有机螯合剂,改善铁的吸收,促进根和芽中FeIII-螯合还原酶的活性,从而增加叶子中铁的浓度和光合作用活性,特别是在幼苗中(Cerdán等人,2013;Jeong等人,2008;Olanrewaju等人,2019)。事实上,V-PHs的使用已被证明能够改善罗勒、菠菜和多年生壁虎叶片的铁(和一般离子)含量、颜色状态、光合作用、产量和质量(Carillo等人,2019b;Caruso等人,2019;Rouphael等人,2021)。所有这些由V-PHs施加的有益影响可能被描述为 "养分获取反应",并导致养分的吸收、转运和同化的普遍改善(Carillo等人,2019b;Cristofano等人,2022;Sestili等人,2018)。
不可否认的是,这两种类型的生物刺激剂都是对农业实践的有效补充,能够提高作物产量和农产品质量。然而,虽然V-PHs没有不良影响,但有必要密切关注A-PHs的剂量,以避免代谢和生长的负面障碍,并优化作物反应(图1)。
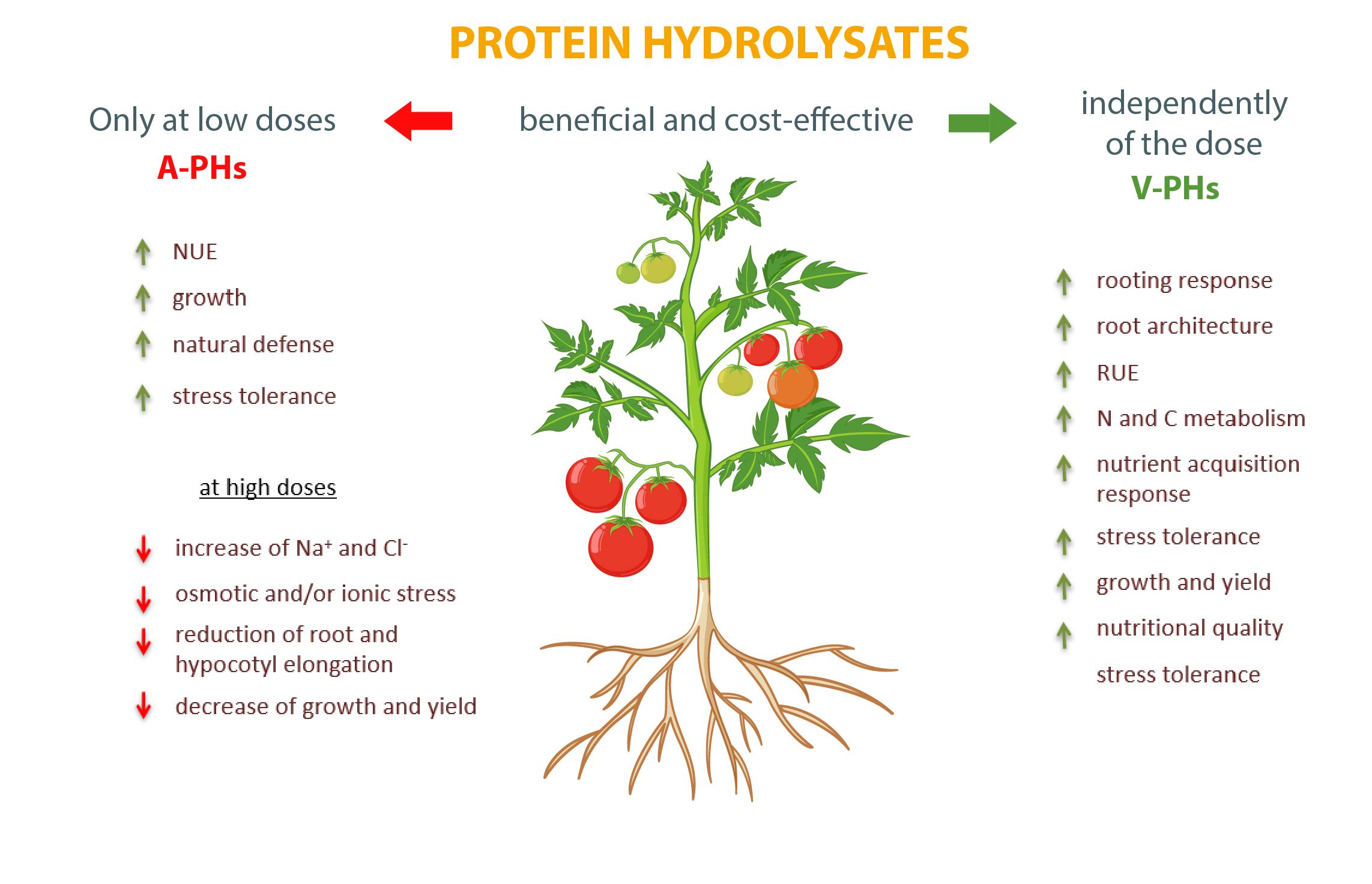
图1. 动物和植物蛋白水解物(A-PHs和V-PHs)对蔬菜作物产生的主要剂量依赖效应。

