
植物作为生态系统的初级生产者,承载着无数的微生物物种,它们依靠富含碳元素、根部排出的物质获得营养,包括糖类、有机酸和氨基酸。真菌和细菌微生物群的很大一部分保持中立,但少数物种可能建立病原体或共生关系,最终影响植物的适应性、适应性和生产力(1)。实现可持续作物管理的一个主要目标是识别和描述具有益生菌特性的微生物物种,从中可能发现高活性物质,为不断增长的市场开发新一代的农药、生物刺激剂和防御诱导剂。在这种情况下,真菌属的重要性日益增加,因为它包括200多个报道的物种,具有多功能的代谢和良好的适应性,可以在土壤和水环境中增殖(2)。
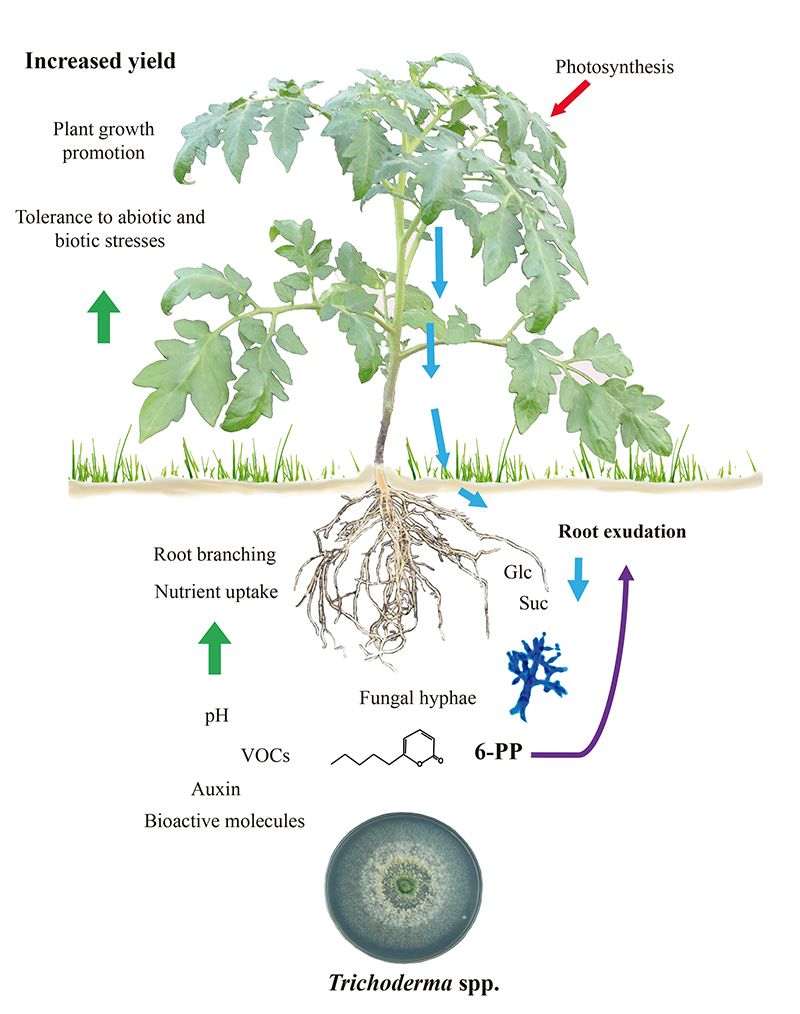
图1.
毛霉菌-诱导的植物生长和防御取决于根部渗出物
植物的生长/防御权衡在生物相互作用中被调节。这确保了发育过渡到开花的过程,并保证种子生产。由于毛霉菌同时触发了植物的免疫和生长促进,很难确定这些固有的代价的程序是否可能有联系。此外,真菌定居的根部可能有一个增强的新陈代谢,以支持共生体的能量需求。已经发现番茄幼苗的根部渗出的碳水化合物在对阿特罗维德菌的反应中有所增加,糖类可能起到积极的趋化反应,以吸引菌丝(11),如图1中的报告。一致的是,该真菌表达一个细胞内的转化酶(Triat51014)和两个推定的蔗糖运输器(Triat226844和Triat83012)来使用植物来源的碳。另一方面,T. virens通过蔗糖转运体吸收蔗糖,并通过细胞内的转化酶进行水解,通过这种二糖提供的能量使其快速生长(12)。
分子证据有助于解释毛霉菌如何在检测到根部渗出物后改变其代谢特征。Villalobos-Escobedo等人(2020)进一步证明,在T. atroviride突变体中,编码参与复杂碳水化合物降解(如纤维素或甲壳素)的基因的表达受到影响,NADPH氧化酶是一种直接参与生产活性氧的酶。相应基因的突变不仅影响植物宿主的侧根形成和生物量生产,而且还影响引起的防御反应和真菌的滋生行为,以获得和利用根部渗出物中的简单糖类(3)。
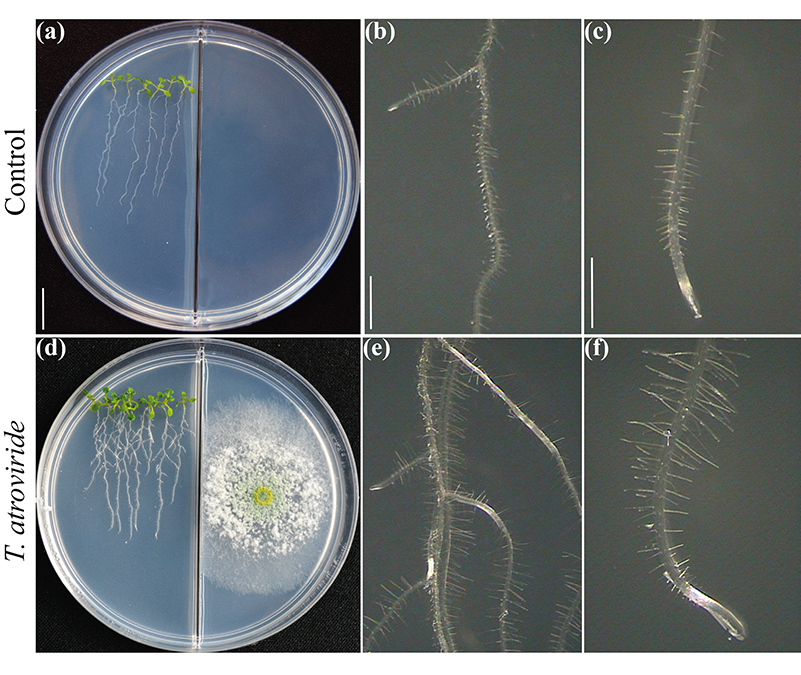
图2.
人们在很大程度上讨论了在与微生物的相互作用中,作为高生物量生产基础的植物性状是否主要依赖于排放的二氧化碳。当然,二氧化碳是光合作用的主要反应物,应该是生物量提高的原因。然而,紧密的交流取决于挥发性混合物的分子组成。通过在分割的培养皿中测试植物的生长和发育,评估真菌与植物通过挥发物的相互作用(图2),我们已经揭示了6-PP对植物生物量生产的关键作用,它直接影响激素反应,以及芽-根长距离运输糖类(Esparza-Reynoso等人提交)。它支持这样的观点,即取决于光合作用、嫩枝的碳状态和真菌代谢产物的感知的糖分配调节回路对根分生组织的有丝分裂至关重要。有趣的是,促进植物生长的真菌,包括Trichoderma 菌株和Serendipita indica和S. williamsii ,不是通过单一的化合物,而是通过挥发物的混合物,强烈促进光合作用(13, 14)。在暴露于真菌挥发物的叶片中,显然不能发生由于糖水平升高而导致的二氧化碳固定的反馈抑制,这可能是由于荷尔蒙不平衡。植物并不孤独,它们依靠它们的真菌共生体来生存和发展。现在已经到了将目前的知识转化为实施毛霉菌的田间应用的时候了。
- Cordovez V, Dini-Andreote F, Carrión VJ, Raaijmakers JM(2019)。植物微生物组的生态学和进化。Rev. Microbiol.73:69-88.
- López-Bucio J, Pelagio-Flores R, Herrera-Estrella A(2015)。 作为生物刺激剂的毛霉菌:利用一种植物有益真菌的多层次特性。 Hort. 196:109-123.
- Villalobos-Escobedo JM, Esparza-Reynoso S, Pelagio-Flores R, López-Ramírez F, Ruiz-Herrera LF, López-Bucio J, Herrera-Estrella A(2020)。真菌NADPH氧化酶是毛霉菌 和拟南芥之间分子对话的一个重要元素。Plant J. 103:2178-2192.
- Garnica-Vergara A, Barrera-Ortiz S, Muñoz-Parra E, Raya-González J, Méndez-Bravo A, Macías-Rodríguez L, Ruiz-Herrera LF, López-Bucio J(2016)。来自毛霉菌 的挥发性6-戊基-2H-吡喃-2-酮通过辅助素信号和ETHENE INSENSITIVE 2功能调节拟南芥根部形态发生。New Phytol.209:1496-1512.
- Lee S, Behringer G, Hung R, Bennett J (2019).真菌挥发性有机化合物对拟南芥 生长和基因表达的影响。Fungal Ecol. 37:1-9.
- Guo Y, Ghirardo A, Weber B, Schnitzler JP, Benz JP, Rosenkranz M(2019)。 Trichoderma物种在其挥发性概况和对外生菌株 Laccaria bicolor的拮抗作用方面存在差异。 Microbiol. 10:891.
- Pelagio-Flores R, Esparza-Reynoso S, Garnica-Vergara A, López-Bucio J, Herrera-Estrella A(2017)。毛霉菌诱导的酸化是拟南芥根部生长变化的早期触发器,决定了真菌的植物刺激。 Plant Sci.8:822.
- Contreras-Cornejo HA, Macías-Rodríguez L, Cortés-Penagos C, López-Bucio J(2009)。毛霉菌,一种植物有益的真菌,在拟南芥中通过辅助素依赖性机制提高生物量生产并促进侧根生长。植物生理学》,149:1579-1592。
- Wang YF, Hou XY, Deng JJ, Yao ZH, Lyu MM, Zhang RS (2020).AUXIN RESPONSE FACTOR 1在过量表达的杨树对Trichoderma asperellum接种的反应中作为一个正向调节器。植物(巴塞尔)9:272。
- Rocafort M, Fudal I, Mesarich CH (2020).植物相关真菌和卵菌的细胞膜效应蛋白。 观点。 56:9-19 。
- Macías-Rodríguez L, Guzmán-Gómez A, García-Juárez P, Contreras-Cornejo HA(2018)。 阿特罗维德毛霉菌促进了番茄的发育,改变了根部渗出的碳水化合物,从而刺激了真菌的生长和三方互动系统中植物病原体Phytophthora cinnamomi的生物控制。FEMS Microbiol.Ecol. 94:fiy137.
- Vargas WA, Mandawe JC, Kenerley CM (2009). 植物来源的蔗糖是毛霉菌 与玉米植物共生关系中的一个关键因素。植物生理学 》,151:792-808。
- Harman GE, Doni F, Khadka RB, Uphoff N(2019)。毛霉菌的内生菌株提高了植物的光合作用能力。 Appl. Microbiol. https://doi.org/10.1111/jam.14368.
- Venneman J. et al. (2020).呼吸性二氧化碳与内生性Serendipita菌株排放的混合挥发物相结合,强烈刺激拟南芥的生长,牵涉到辅助素和细胞分裂素信号的传递。 Plant Sci. https://doi.org/10.3389/fpls.2020.544435.

