
Растения как первичные производители экосистем являются хозяевами огромного количества видов микроорганизмов, которые для своего питания используют богатые углеродом вещества, выделяемые корнями, включая сахара, органические кислоты и аминокислоты. Большая часть грибкового и бактериального микробиома остается нейтральной, но некоторые виды могут устанавливать патогенные или симбиотические отношения, которые в конечном итоге влияют на приспособленность, адаптацию и продуктивность растений (1). Основной целью устойчивого управления сельскохозяйственными культурами является выявление и характеристика микробных видов с пробиотическими свойствами, из которых можно выделить высокоактивные вещества для разработки нового поколения пестицидов, биостимуляторов и защитных элиситоров для растущего рынка. В этом сценарии все большее значение приобретает род грибов Trichoderma, который включает более 200 видов с разнообразным метаболизмом и хорошо приспособленными свойствами для размножения в почве и водной среде (2).
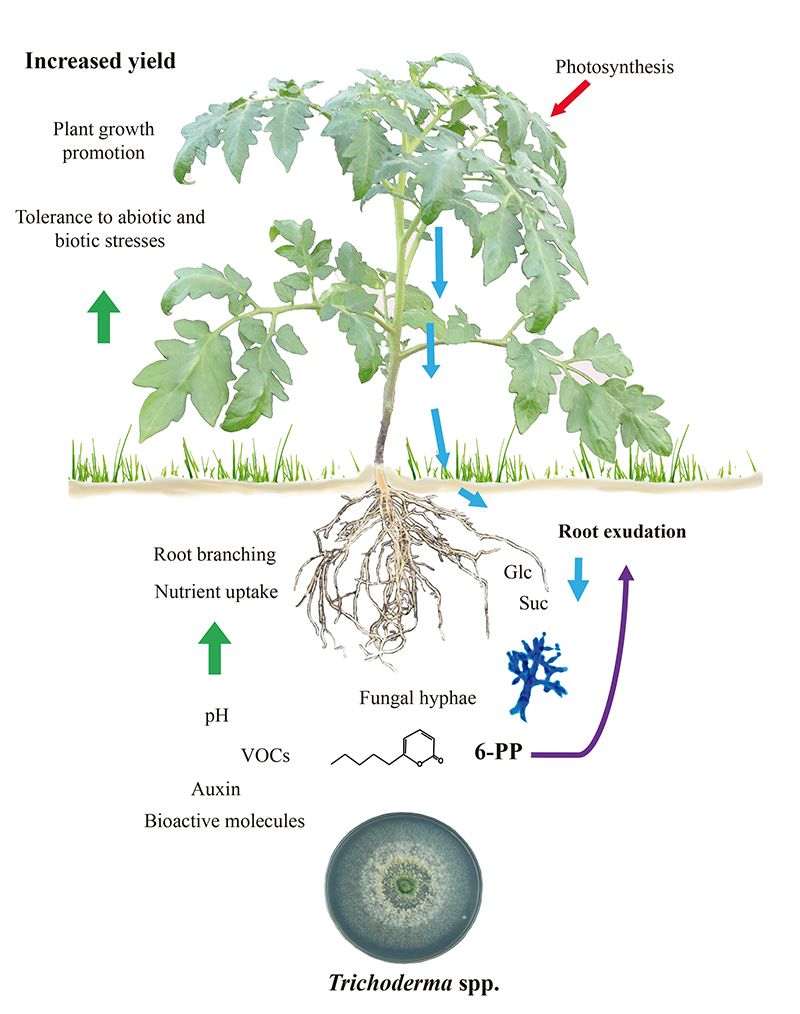
Рисунок 1.
Trichoderma-индуцированный рост и защита растений зависят от корневых экссудатов
Компромиссы между ростом и защитой у растений модулируются во время биотических взаимодействий. Это обеспечивает переход развития к цветению и гарантирует производство семян. Поскольку Trichoderma вызывает как иммунитет растений, так и стимулирование роста, трудно установить, связаны ли эти программы, которые по своей природе являются дорогостоящими. Более того, возможно, что колонизированные грибами корни могут иметь усиленный метаболизм для поддержания энергетических потребностей симбиоза. Было обнаружено, что экссудация углеводов корнями проростков томата увеличивается в ответ на T. atroviride, и сахара могут действовать как положительный хемотаксический ответ для привлечения гиф (11), как показано на рисунке 1. Последовательно, гриб экспрессирует внутриклеточную инвертазу (Triat51014) и два предполагаемых транспортера сахарозы (Triat226844 и Triat83012) для использования углерода, полученного из растений. С другой стороны, T. virens поглощает сахарозу через транспортер сахарозы и гидролизует ее с помощью внутриклеточной инвертазы, что обеспечивает быстрый рост за счет энергии, предоставляемой этим дисахаридом (12).
Молекулярные данные помогают объяснить, как Trichoderma изменяет свои метаболические сигнатуры при обнаружении корневых экссудатов. Villalobos-Escobedo et al. (2020) пошли дальше и продемонстрировали, что экспрессия генов, кодирующих ферменты, участвующие в деградации сложных углеводов, таких как целлюлоза или хитин, перед колонизацией корней нарушается у мутантов T. atroviride, дефектных по NADPH-оксидазе, ферменту, непосредственно участвующему в производстве реактивных форм кислорода. Мутация соответствующих генов не только влияет на образование боковых корней и производство биомассы в растении-хозяине, но также влияет на вызываемые защитные реакции и сапрофитное поведение гриба для получения и использования простых сахаров, доступных в корневых экссудатах (3).
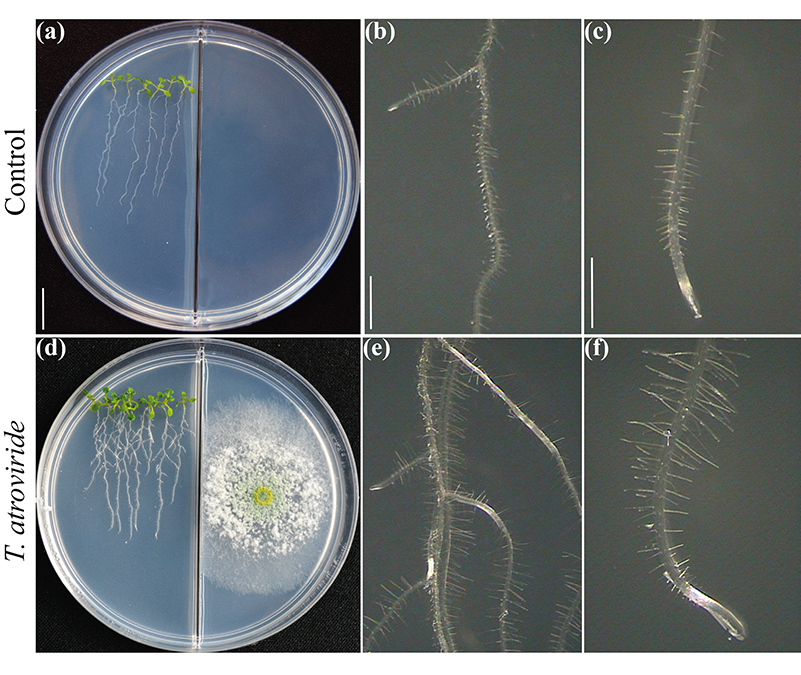
Рисунок 2.
В основном обсуждалось, зависят ли признаки растений, лежащие в основе высокого производства биомассы при взаимодействии с микробами, в основном от выделяемого углекислого газа. Конечно, углекислый газ является основным реактивом для фотосинтеза и должен обеспечивать увеличение биомассы. Тем не менее, тесная связь зависит от молекулярного состава летучих смесей. Тестируя рост и развитие растений в разделенных пластинах Петри для оценки взаимодействия грибов и растений через летучие вещества (рис. 2), мы выявили критическую роль 6-PP для производства биомассы растений, непосредственно влияя на гормональные реакции и перенос сахаров на большие расстояния между побегом и корнем (Esparza-Reynoso et al. submitted). Это поддерживает идею о том, что регуляторная петля для распределения сахаров в зависимости от фотосинтеза, углеродного статуса побега и восприятия грибковых метаболитов является критической для митоза в корневых меристемах. Интересно, что грибы, способствующие росту растений, включая штаммы Trichoderma , Serendipita indica и S. williamsii , сильно стимулируют фотосинтез не с помощью отдельных соединений, а через смеси летучих веществ (13, 14). Ингибирование фиксации углекислого газа, вызванное повышением уровня сахара, по-видимому, не происходит в листьях, подверженных воздействию грибных летучих веществ, возможно, из-за гормонального дисбаланса. Растения не одиноки, они полагаются на своих грибковых симбионтов для выживания и процветания. Настало время перевести текущие знания в применение Trichoderma для полевого использования.
- Cordovez V, Dini-Andreote F, Carrión VJ, Raaijmakers JM (2019). Экология и эволюция растительных микробиомов. Rev. Microbiol. 73:69-88.
- López-Bucio J, Pelagio-Flores R, Herrera-Estrella A (2015). Trichoderma как биостимулятор: использование многоуровневых свойств полезного для растений гриба. Hort. 196:109-123.
- Villalobos-Escobedo JM, Esparza-Reynoso S, Pelagio-Flores R, López-Ramírez F, Ruiz-Herrera LF, López-Bucio J, Herrera-Estrella A (2020). Грибковая NADPH-оксидаза - важный элемент для молекулярного диалога между Trichoderma и Arabidopsis. Plant J. 103:2178-2192.
- Garnica-Vergara A, Barrera-Ortiz S, Muñoz-Parra E, Raya-González J, Méndez-Bravo A, Macías-Rodríguez L, Ruiz-Herrera LF, López-Bucio J (2016). Летучий 6-пентил-2H-пиран-2-он из Trichoderma atroviride регулирует корневой морфогенез Arabidopsis thaliana через сигнализацию ауксина и функционирование ETHYLENE INSENSITIVE 2. New Phytol. 209:1496-1512.
- Lee S, Behringer G, Hung R, Bennett J (2019). Влияние летучих органических соединений грибов на рост и экспрессию генов Arabidopsis thaliana . Fungal Ecol. 37:1-9.
- Guo Y, Ghirardo A, Weber B, Schnitzler JP, Benz JP, Rosenkranz M (2019). Виды Trichoderma различаются по своим летучим профилям и антагонизму к эктомикоризе Laccaria bicolor. Microbiol. 10:891.
- Pelagio-Flores R, Esparza-Reynoso S, Garnica-Vergara A, López-Bucio J, Herrera-Estrella A (2017). Индуцированное триходермой закисление является ранним триггером изменений в росте корней арабидопсиса и определяет грибковую фитостимуляцию. Plant Sci. 8:822.
- Contreras-Cornejo HA, Macías-Rodríguez L, Cortés-Penagos C, López-Bucio J (2009). Trichoderma virens, полезный для растений гриб, увеличивает производство биомассы и способствует росту боковых корней через ауксин-зависимый механизм в Arabidopsis. Физиол растений. 149:1579-1592.
- Wang YF, Hou XY, Deng JJ, Yao ZH, Lyu MM, Zhang RS (2020). AUXIN RESPONSE FACTOR 1 действует как положительный регулятор в ответе тополя на инокуляцию Trichoderma asperellum в сверхэкспрессирующих растениях. Plants (Basel) 9:272.
- Rocafort M, Fudal I, Mesarich CH (2020). Апопластические эффекторные белки растительно-ассоциированных грибов и оомицетов. Opin. Plant Biol. 56:9-19.
- Macías-Rodríguez L, Guzmán-Gómez A, García-Juárez P, Contreras-Cornejo HA (2018). Trichoderma atroviride способствует развитию томата и изменяет корневую экссудацию углеводов, что стимулирует рост грибов и биоконтроль фитопатогена Phytophthora cinnamomi в системе трехстороннего взаимодействия. FEMS Microbiol. Ecol. 94:fiy137.
- Vargas WA, Mandawe JC, Kenerley CM (2009). Полученная из растений сахароза является ключевым элементом симбиотической ассоциации между Trichoderma virens и растениями кукурузы. Физиол растений. 151:792-808.
- Harman GE, Doni F, Khadka RB, Uphoff N (2019). Эндофитные штаммы Trichoderma повышают способность растений к фотосинтезу. Appl. Microbiol. https://doi.org/10.1111/jam.14368.
- Venneman J. et al. (2020). ДыхательныйCO2 в сочетании со смесью летучих веществ, выделяемых эндофитными штаммами Serendipita, сильно стимулирует рост арабидопсиса, связывая ауксин и цитокининовую сигнализацию. Plant Sci. https://doi.org/10.3389/fpls.2020.544435.


© BIOSTIMULANT.COM
ВСЕ ПРАВА ЗАЩИЩЕНЫ