
Bien que de nombreuses publications confirment les avantages liés à l'utilisation des biostimulants, les mécanismes moléculaires qui sous-tendent ces effets positifs sont encore mal élucidés. Néanmoins, la compréhension du mode d'action par lequel les biostimulants exercent leur activité peut fournir des indications utiles pour mieux définir la ou les cibles en termes de cultures, d'allégations, de pratiques agricoles et de calendrier d'application. De plus, la récente réglementation adoptée par l'UE suggère l'opportunité de comprendre les processus de culture affectés par le biostimulant afin de soutenir les allégations souhaitées au niveau réglementaire.
Partant de ce principe, nous pouvons souligner que répondre à la demande susmentionnée constitue un défi crucial. Cela est dû, principalement, à deux raisons : (i) la complexité des processus biochimiques des plantes, dont certains ne sont pas encore totalement élucidés ; (ii) la variabilité de la réponse génotypique, plutôt que de cultivar à cultivar, et le rôle central de modélisation exercé par l'environnement et les pratiques agronomiques.
Ancrées dans le sol, les plantes ont dû développer une grande diversité de processus biochimiques pour répondre aux stress abiotiques et biotiques ; les biostimulants végétaux ciblent/modulent souvent ces processus. Plusieurs approches peuvent être utilisées pour faire la lumière sur les variations biochimiques imposées par des facteurs externes tels que l'utilisation de biostimulants sur les cultures. Parmi elles, les approches non ciblées sont à recommander car elles ne nécessitent pas d'hypothèses a priori, se prêtant ainsi à des scénarios complexes. Les sciences omiques (par exemple, la génomique, la transcriptomique, la protéomique et la métabolomique) se rapportent au profil des gènes, des transcriptions, des protéines et des métabolites dans un système biologique et relèvent des approches non ciblées. Parmi les différentes omiques, la métabolomique est la plus proche du génotype et a été efficacement proposée pour étudier le mode d'action des biostimulants végétaux. En effet, la modulation de l'expression des gènes s'accompagne d'une modification post-traductionnelle des protéines, qui se traduit finalement par le profil actuel des métabolites. Les progrès récents en matière d'équipement analytique, comme la spectrométrie de masse à haute résolution, la disponibilité des bases de données et la gestion des données (y compris l'analyse statistique multivariée et les outils bioinformatiques pour l'interprétation des données) ont stimulé l'adoption de la métabolomique dans les sciences agricoles. En conséquence, des différences dans les signatures métabolomiques ont été proposées pour identifier les processus végétaux affectés par des facteurs externes tels que les stress abiotiques (salinité, sécheresse, inondation, températures élevées, etc.), les interactions plantes-microbes et plantes-pathogènes, et les biostimulants (Fig. 1).
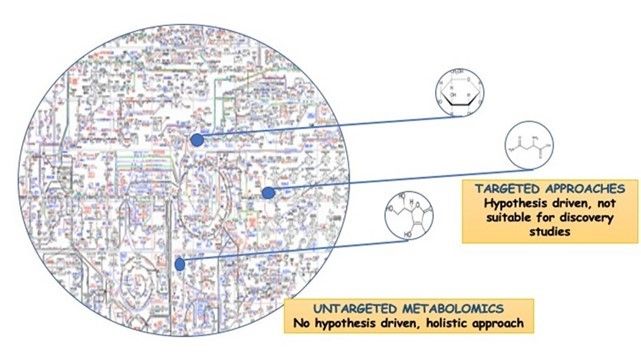
Figure 1. La métabolomique non ciblée est l'analyse complète de tous les métabolites mesurables dans un échantillon, y compris les inconnus chimiques, et la métabolomique ciblée, la mesure de groupes définis de métabolites chimiquement caractérisés et biochimiquement annotés.
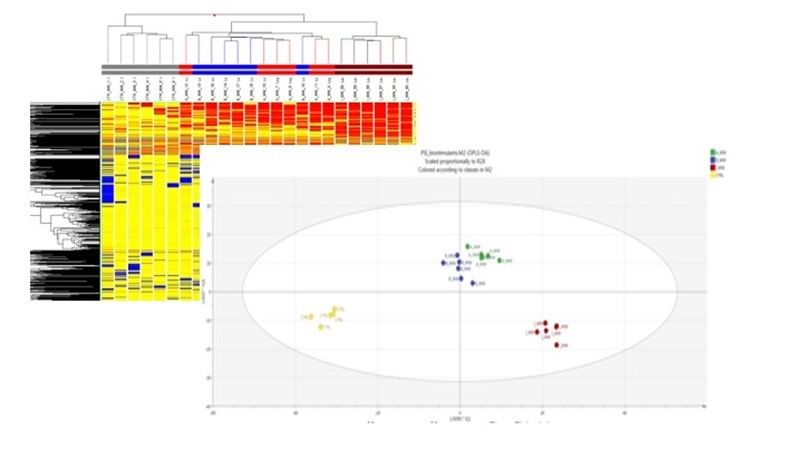
Figure 2. Analyse de regroupement hiérarchique non supervisée réalisée à partir de profils métabolomiques après application d'hydrolysats de protéines sélectionnés (figure supérieure). Graphique de score de la modélisation supervisée de l'analyse discriminante par projection orthogonale aux structures latentes (OPLS-DA) effectuée sur des profils métabolomiques après application d'hydrolysats de protéines sélectionnés, où la variation entre les groupes a été séparée en composantes prédictives et orthogonales (figure du bas).
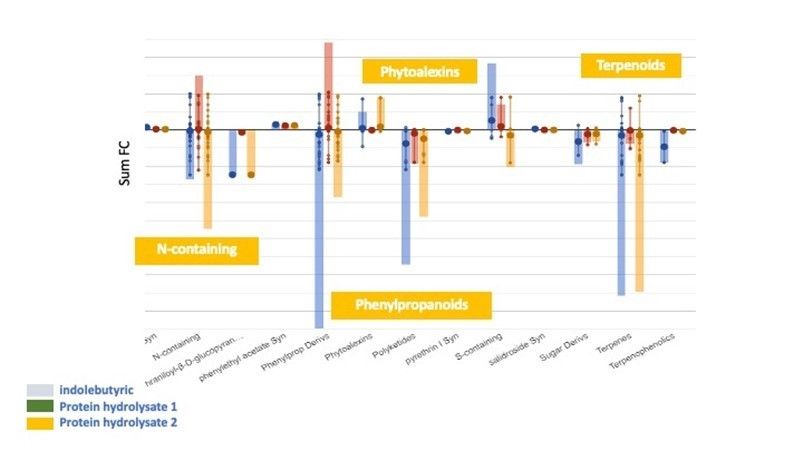
Figure 3. Processus de biosynthèse impliqués dans les boutures de tomates à l'application foliaire d'hydrolysats de protéines d'origine végétale et à l'application sur la tige d'acide indole-butyrique (IBA) en utilisant une immersion rapide basale dans une solution concentrée d'IBA. L'axe des abscisses représente chaque ensemble de sous-catégories tandis que l'axe des ordonnées correspond à la variation en plis cumulée par rapport au témoin non traité.


©2020 BIOSTIMULANT.COM
TOUS DROITS RÉSERVÉS