
Nitrogen is the primary nutrient required for plant growth in the largest quantities and thus essential for agricultural productivity. The global application of synthetic nitrogen fertilizer has increased by 10-fold from 1950 to 2008 (Robertson and Vitousek, 2009). The USDA reports a 5-time increase in the consumption of nitrogen fertilizer from 1960 to 2014, during when the average U.S. farm price of nitrogen-sourced fertilizers such as nitrate, ammonium, urea or other organic forms increased by more than 30 times (USDA ERS, 2019) (Fig. 1). It is critical to accommodate rapidly growing global food demands while effectively addressing concerns over excessive agrochemical use for sustainable food production.
Fertilizer is a relatively small portion of total production cost, and therefore, excess fertilizers are often applied to provide sufficient plant nutrients and ensure high crop yield. However, this management practice can lead to environmental impacts, including contamination of water bodies resulting from leaching and runoff from the production sites and an increase in the risk of global warming from the release of nitrous oxide (N2O), a potent greenhouse gas. Agriculture is the largest source of N2O emissions in the United States derived from nitrogen fertilization, which accounts for nearly 77.8 percent of total N2O emissions in 2018 (US EPA, 2015). Further, excess nitrogen fertilization can produce undesirable plants with lush growth that are susceptible to disease and promote nitrate accumulation in edible parts of the plants.
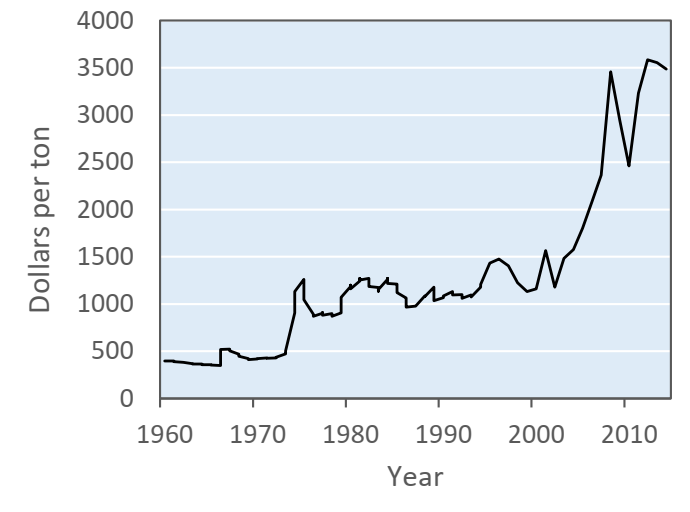
Fig. 1. Increase in total U.S. farm price of nitrogen fertilizers from 1960 to 2014. The data are the sum of average U.S. prices of selected fertilizers such as anhydrous ammonia, nitrogen solutions (30%), urea (44-46% nitrogen), ammonium nitrate, sulfate of ammonium, and diammonium phosphate (18-46-0).
Especially, nitrate accumulation in the edible portions of crops is a major concern to human health. According to the recommendations of the FAO/WHO Expert Committee on Food Additives (JECFA, 2002), the daily intake of nitrates by an individual should not exceed 3.7 mg/kg body weight. Excessive dietary intake of nitrate and its derivatives such as nitrite, nitric oxide, and N-nitroso compounds, are associated with a serious type of anemia called methemoglobinemia in young infants and a higher risk of human health problem such as gastric and bladder cancers (Colla et al., 2018). A known strategy for minimizing nitrate accumulation in crops is to withhold or reduce nitrogen input before harvest. This practice is effective especially in leafy vegetables because nitrogen requirements of crops decrease as plant growth matures and forces stored nitrates to be used to support growth, allowing plants to accumulate organic compounds in place of the declining osmoticum. An alternative and promising strategy is to apply plant biostimulants in combination with nitrogen fertilization to prevent crops from accumulating excess nitrogen while avoiding the risk of compromising crop yield and quality.
Plant biostimulants have received significant attention in recent years as a technology innovation for the sustainability of agriculture systems. Plant biostimulants are a large group of natural substances and microbial inoculants, including protein hydrolysates (PH), seaweed extracts (SE), humic substances (HS), arbuscular mycorrhizal fungi (AMF), and plant growth-promoting bacteria (PGPB). These biostimulants are proven to improve crop growth and yield, nutritional quality, and abiotic and biotic stress tolerance with only a small amount of application (Colla and Rouphael, 2015). Plant biostimulants also enhance soil structure by simulating microbial activities and improve plant nitrogen uptake by altering plant morphological and physiological characteristics (Halpern et al., 2015) (Fig. 2). One of the well-known effects of plant biostimulants is a marked increase in root growth. The changes in the root architecture allow plants to explore a larger volume of soil and effectively capture nutrients. These functions are considered stemmed from the presence of bioactive peptides inducing hormone-like activities (auxin, gibberellins, and brassinosteroids), which involve complex interactions among phytohormones (Colla et al., 2015; Kim et al., 2019). Several studies also suggest that plant biostimulants promote nitrogen reduction and assimilation through coordinate regulation of nitrogen metabolic pathways (Schiavon et al, 2008; Sestili et al., 2018).
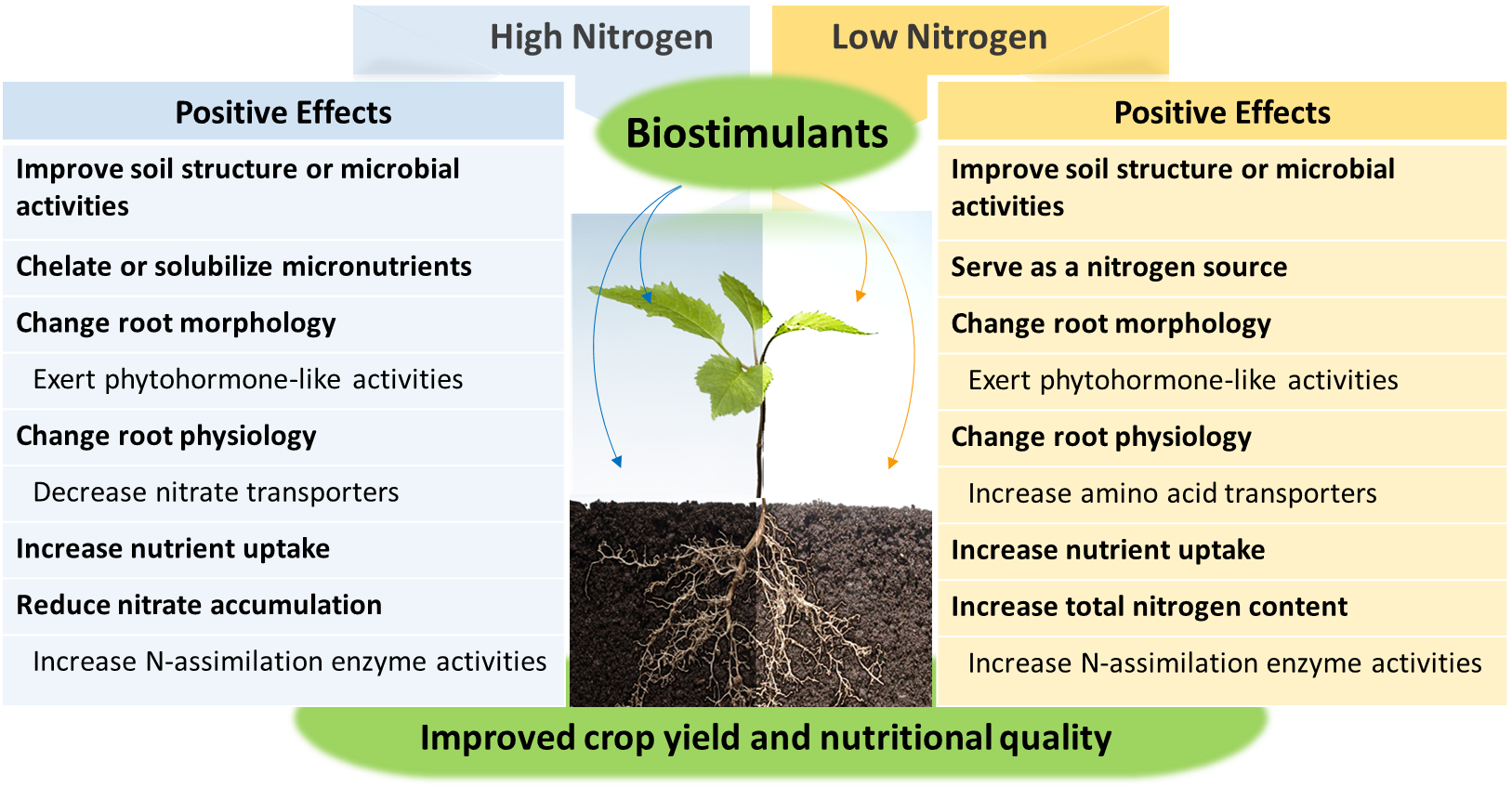
Fig. 2. The positive effects of biostimulants on crop yield and quality under high (left) and low (right) nitrogen conditions.
Another well-known effect of plant biostimulants is the yield and quality improvement of the crops. Differential mechanisms are involved in plant responses to the biostimulants depending on nitrogen fertilization conditions (Fig. 2). Biostimulants can safeguard plants from low nitrogen by improving crop yield and quality and from high nitrogen by reducing nitrate accumulation. The latter effects are not unequivocal and largely depend on the crop, environmental conditions, and the type of biostimulants applied, but the weight of the evidence supports this contention. For example, PH decreased nitrates in leafy vegetables including rocket, Swiss chard, spinach, lettuce, and parsley, as well as celery and radish, under high nitrogen (Colla et al., 2015). Likewise, the application of a PH solution reduced nitrate accumulation in lettuce grown under high nitrogen (Tsouvaltzis et al., 2014). PH did not affect nitrate content in perennial wall rocket (Giordano et al., 2020) but increased it in the leaves of spinach (Carillo et al., 2019). An increase of nitrate accumulation was observed in rocket applied with PH, but it was maintained under the legal limits set by the European Commission (Di Mola et al., 2019a).
The effects of plant biostimulants on nitrate accumulation in crops varied by the types of synthetic nitrogen fertilizers used or climactic conditions, indicating the interaction between plant biostimulants and environmental factors. In a study where various nitrogen fertilizers were evaluated in combination with AMF (Glomus intraradices), nitrate accumulation in lettuce was reduced or not affected when inoculated with AMF but increased when urea was used as the source of nitrogen (Mitova et al., 2017). The beneficial effect of PGPB (Enterobacter sp. and Bacillus sp.) inoculation on transplants disappeared during field production due to low light intensity and temperatures in the late season, and nitrate content in the leaves of lettuce increased regardless of PGPB inoculation (Szczech et al., 2016). These findings suggest that environmental factors have significant impacts on nitrate accumulation in crops and can mask the beneficial effects of plant biostimulants. The interactions of biostimulants and environmental factors need further investigations to clarify this aspect.
Meanwhile, biostimulants serve as an organic nitrogen source (i.e., amino acids and peptides) under limited nitrogen conditions and improve the growth and yield and nutritional quality of crops by increasing total nitrogen of the crops (Fig. 2). A recent study showed that such effects induced by PH were associated with the upregulation of the expression of genes encoding amino acid transport and transcripts of the enzymes involved in nitrogen assimilation (Sestili et al., 2018). Foliar application of PH increased the yield and total amino acid content in greenhouse spinach under low nitrogen (Carillo et al., 2019). Similarly, foliar spray of SE improved the yield of baby lettuce under low nitrogen; however, leaf nitrate content also increased but to a level below the legal limits (Di Mola et al., 2019b). Inoculation with either AMF or PGPB increased total nitrogen in the shoots of strawberry plants under reduced fertilization with some variations in the effect by the strains (Lingua et al., 2013). A combined inoculation of plants with PGPB and AMF improved tomato fruit yield and nutrition (β-carotene) under reduced fertilization (Bona et al., 2018). A similar observation was made in lettuce plants exposed to water stress, where the inoculation with PGPB, alone or in combination with AMF, stimulated nitrate reductase activity in the leaves, increasing total nitrogen content (Kohler et al., 2008). A biostimulant mixture containing PH and HS maintained fruit yield and quality parameters under reduced fertilization while increasing total soluble protein in tomato fruits (Koleška et al., 2017). Collective information from these studies provides insights into the function of plant biostimulants in improving the yield and nutritional quality of crops when nitrogen is limited.
Current knowledge provides promising snapshots on the use of biostimulants for agricultural production. Overall, the application of biostimulants protects crops from potential risks of yield and quality loss and helps produce crops safely and sustainably. Given that plant biostimulants are used in combination with reduced nitrogen fertilizer, this approach has the potential to improve the nitrogen use efficiency of the production systems. In light of many benefits that can bring to crop production systems, plant biostimulants provide a viable option for farmers, addressing food security, nutrition, and environmental sustainability.
Bona, E., Todeschini, V., Cantamessa, S., Cesaro, P., Copetta, A., Lingua, G., Gamalero, E., Berta, G., Massa, N. 2018. Combined bacterial and mycorrhizal inocula improve tomato quality at reduced fertilization. Sci. Hortic. 234, 160–165
Carillo, P., Colla, G., Fusco G.M., Aversana, E.D., El-Nakhel, C., Giordano, M., Pannico, A., Cozzolino, E., Mori, M., Reynaud, H., Kyriacou, M.C., Cardarelli, M., Rouphael, Y. 2020. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 9 (8), 450
Colla, G., Kim, H.J., Kyriacous, M.C. and Rouphael. Y. 2018. Nitrate in fruits and vegetables. Sci. Hortic. 237: 221-238
Colla, G., Nardi, S., Cardarelli, M., Ertani, A., Lucini, L., Canaguier, R., Rouphael, Y. 2015. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 96, 28–38
Colla, G., Rouphael, Y. 2015. Biostimulants in horticulture. Sci. Hortic. 196, 1–2
Di Mola, I., Cozzolino, E., Ottaiano, L., Giordano, M., Rouphael, Y., Colla, G., Mori, M., 2019a. Effect of vegetal- and seaweed extract-based biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 9(10), 571
Di Mola, I.D., Ottaiano, L., Cozzolino, E., Senatore, M., Giordano, M., El-Nakhel, C., Sacco, A., Rouphael, Y., Colla, G., Mori, M. 2019b. Plant-based biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 8, 522
Giordano, M., El-Nakhel, C., Caruso, G., Cozzolino, E., Pascale, S.D, Kyriacou, M.C., Colla, G., Rouphael, Y. 2020. Stand-alone and combinatorial effects of plant-based biostimulants on the production and leaf quality of perennial wall rocket. Plants 9, 922
Halpern, M., Bar-Tal, A., Ofek, M., Minz, D., Muller, T., Yermiyahu, U. 2015. The use of biostimulants for enhancing nutrient uptake, in: Advances in Agronomy. Elsevier, pp. 141–174
JECFA. 2002. Joint FAO/WHO Expert Committee on Food Additives, Evaluation of certain food additives and contaminants. Fifty-ninth report of the Joint FAO/WHO Experts Committee on Food Additives. WHO Technical Reports series 913. 20–32
Kim, H.J., Ku, K.M., Choi, S.H., Cardarelli. M. 2019. Biostimulant enhances adventitious rooting in cuttings of basil, tomato, and chrysanthemum via brassinosteroid-mediated processes. Agronomy 9(2): 74
Kohler, J., Hernández, J.A., Carabaca, F., Roldán, A. 2008. Plant growth promoting rhizobacteria and arbuscular mycorrhizal fungi modify alleviation biochemical mechanism in water stressed plants. Funct. Plant Biol. 35, 141–151
Koleška, I., Hasanagić, D., Todorović, V., Murtić, S., Klokić, I., Parađiković, N., Kukavica, B., 2017. Biostimulant prevents yield loss and reduces oxidative damage in tomato plants grown on reduced NPK nutrition. J. Plant Interact. 12, 209–218
Lingua, G., Bona, E., Manassero, P., Marsano, F., Todeschini, V., Cantamessa, S., Copetta, A., D’Agostino, G., Gamalero, E., Berta, G., 2013. Arbuscular mycorrhizal fungi and plant growth-promoting Pseudomonads increases anthocyanin concentration in strawberry fruits (Fragaria ´ ananassa var. Selva) in conditions of reduced fertilization. Int. J. Mol. Sci. 14, 16207–16225
Mitova, I., Nenova, L., Stancheva, I., Geneva, M., Hristozkova, M., Mincheva, J. 2017. Lettuce response to nitrogen fertilizers and root mycorrhization. Bulgarian J. Agric. Sci. 23, 260–264
Robertson, G.P., Vitousek, P.M., 2009. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 34, 97–125
Schiavon, M., Ertani, A, Nardi, S. 2008. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) cycle and nitrogen metabolism in Zea mays L. J. Agric. Food Chem. 56:11800–11808
Sestili, F., Rouphael, Y., Cardarelli, M., Pucci, A., Bonini, P., Canaguier, R., Colla, G. 2018. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 9:1233
Szczech, M., Szafirowska, A., Kowalczyk, W., Szwejda-Grzybowska, J., Włodarek, A., Maciorowski, R. 2016. The effect of plant growth promoting bacteria on transplants growth and lettuce yield in organic production. J. Horti. Research. 24(2):101–107
Tsouvaltzis, P., Koukounaras, A., Siomos, S.A. 2014. Application of amino acids improves lettuce crop uniformity and inhibits nitrate accumulation induced by the supplemental inorganic nitrogen fertilization. Int. J. Agric. Biol. 16, 951–955
US EPA, 2015. Overview of greenhouse gases. US EPA. https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed 4.27.2020).
USDA ERS – Fertilizer use and price, 2019. https://www.ers.usda.gov/data-products/fertilizer-use-and-price.aspx (accessed 4.27.2020)


© BIOSTIMULANT.COM
ALL RIGHTS RESERVED